Chapter 98 Pathophysiology of Ocular Trauma
Introduction
Ocular trauma is a major cause of ocular morbidity and a leading cause of monocular visual loss. It is estimated that over 2 million eye injuries occur in the USA each year.1 Children and young adults are particularly at risk, and consequently, society suffers significant socioeconomic loss, as well as human loss, from these common injuries. In 1990, national projections estimated that more than 227 000 hospital days and US$175–200 million in hospital charges alone result from ocular injuries every year.2
A classification of ocular trauma with standardized terminology was developed by Kuhn and associates.3 The International Society of Ocular Trauma subsequently used this terminology to develop a classification system for mechanical injuries of the eye (Box 98.1). This system has proved useful for describing ocular trauma without miscommunication and for facilitating the delivery of optimal patient care.
Different types of ocular injuries have different pathophysiologic and therapeutic ramifications; therefore, knowledge of the initial mechanism of injury to the macula or optic nerve is critical for determining visual prognosis.4–6 Subsequent wound healing responses leading to intraocular proliferation, traction retinal detachment and posttraumatic proliferative vitreoretinopathy (PVR) can play major roles in determining the final visual outcome.4,6–9
Before the use of vitrectomy, wound closure often left vitreous incarcerated in the lips of the scleral or corneal edges of the wound, providing a scaffold for future proliferation. In the first large prospective study, conducted between 1952 and 1970, only 6% of patients with open-globe injury gained visual acuity of 5/200 or better.10 Our improved understanding of wound healing and the advent of vitrectomy techniques in the 1970s permitted more successful repair of posterior segment wounds and resulted in a marked decrease in enucleations.8,9,11,12
This chapter reviews the pathophysiology and the therapeutic aspects related to open-globe injury involving the posterior segment. (See also Chapter 91, Traumatic chorioretinopathies; Chapter 97, Pathogenesis of proliferative vitreoretinopathy, and Chapter 110, Surgery for ocular trauma.)
Anatomic change
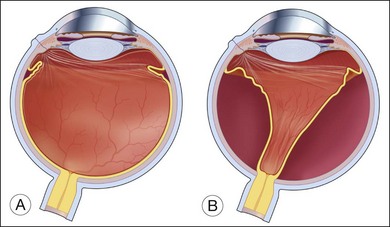
While the direct damage to the ocular tissue depends upon the nature of trauma, the integrity of the vitreoretinal barrier is commonly disrupted by ocular trauma. The vitreous is normally attached to all contiguous structures, including the posterior lens capsule, the pars plana of the ciliary body, the retina, and the optic disc; but the strength of this attachment varies. The vitreous is most firmly attached at its base and is relatively firmly attached to the lens, fovea–parafoveal area, margin of the optic nerve head, and along major retinal blood vessels.13 Weakening of the vitreoretinal interface induced by mechanical force can lead to acute posterior vitreous detachment (PVD) from the retina. When the detachment reaches a point of firmer attachment to the retina, typically the vitreous base, it exerts traction on the retina. The abrupt PVD and/or prolapse and incarceration of the vitreous through the penetrating wound predispose the eye to vitreous traction on the retina and tractional retinal detachment (Fig. 98.1).14–18
Histopathologic findings
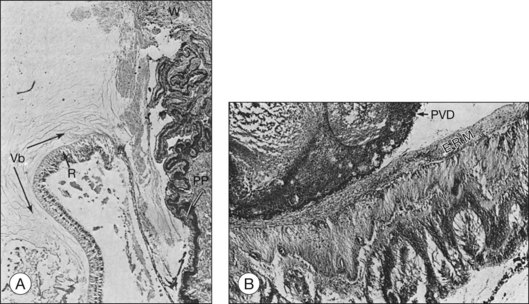
Histopathological evaluation of human eyes enucleated after penetrating trauma has shown that healing of limbal and sclera wounds was more rapid than healing of corneal wounds.17 As early as 4 days after injury, fibroblastic proliferation had occurred from the episclera, and by 1 week, from the stroma of the ciliary body and choroid. At 2 weeks, a mass of vascularized fibrous tissue joined the wound edges; and by 4–6 weeks, a dense fibrous scar had formed. This fibrous ingrowth in limbal or scleral wounds occurred in relation to vitreous incarceration and damage to the lens and/or vitreous hemorrhage. Condensation of vitreous fibrils over the vitreous base was followed by PVD. Typically, the vitreous remained attached anteriorly, and the condensed vitreous fibrils remained attached to the peripheral retina, frequently radiating from a limbal or scleral wound associated with fibroblasts (Fig. 98.2). Retinal detachment was present in most eyes. Although retinal tears were found in a few eyes, it was impossible to exclude a rhegmatogenous component since not all eyes were serially sectioned. Retinal hemorrhage and choroidal hemorrhage were common within the first 2 months and 2 weeks of injury, respectively. Epiretinal membranes were present over both the peripheral and posterior retina by 6 weeks after injury. Subretinal membranes were delicate, branching, and dendritic in appearance 1 and 2 weeks after injury and were thickened and attached to folds in the retina in later phases. Intraocular inflammatory infiltrate, mainly monocytes, was prominent in the anterior chamber or vitreous. Almost all eyes contained some macrophages, either lining the PVD or accumulated in areas of subretinal hemorrhage. Fibroblastic proliferation within the vitreous was present in the area of the wound, resulting in a cyclitic membrane in the early weeks after injury and containing fibroblast-like cells 2 months after injury.
Experimental models
Human specimens obtained from surgery, such as periretinal membranes, vitreous aspirates, and enucleated eyes, provide information about the pathophysiology of open-globe injury.17,18 However, since these specimens often represent only the advanced stages of disease and since they encompass the secondary effects of retinal detachment and PVR, their contributions to our understanding of the complex mechanism of trauma-induced injury are somewhat limited. Therefore, animal models that reproduce various types of ocular trauma have played an important role in our understanding of pathogenesis.
Cleary and Ryan developed penetrating injury models in rabbits and rhesus monkeys using a standard technique.19–24 A knife wound was made using a stab incision through the pars plana and then lengthened to 8 mm with scissors. Vitreous gel prolapsed through the wound and the vitreous face was ruptured in a manner similar to that encountered in the perforated human eye. The wound was then carefully closed and 0.5 cc of autologous blood was injected into the midvitreous. With this standardized method, traction retinal detachment was achieved with remarkable reproducibility. During the second week after injury, the blood changed to a contracted clot and the posterior vitreous detached. As early as 4 weeks after the injury, fibrous tissue grew from the wound into the vitreous, the blood clot formed fibrous tissue, and the posterior vitreous detached. Epiretinal membranes became visible around this time and progressed for up to 15 weeks. The retinal detachment typically occurred between 6 and 11 weeks after the injury. The configuration of the retinal detachment was indicative of the key processes involved. When the vitreous detached posteriorly, the anteroperipheral portion of the vitreous remained firmly attached to the peripheral retina in the area of the vitreous base. Subsequently, the peripheral retina was dragged forward toward the pars plana through its entire circumference, forming a funnel-shaped configuration with full-thickness folds.19,20,25 The presence of intravitreal blood was a potent stimulant to the development of this cascade of the wound healing process. Some 73% of 25 monkey eyes with intravitreal blood injections developed tractional retinal detachments as opposed to only 24% of eyes that received only balanced salt solution injections.19
Penetrating injuries in human eyes are often accompanied by contusions. An animal model used to study this combination employed pigs because pig sclera is sturdy enough to withstand a blunt pellet injury.26–28 An airgun delivered a pellet to the limbus of a pig eye with standardized force at impact. An 8-mm incision was then made in the pars plana as previously described. The main features were the development of intravitreal proliferation and traction retinal detachment. Additionally, subretinal hemorrhage was frequently associated, leading to subretinal fibrous membrane formation.
Animal models are useful in reproducing the findings observed in ocular trauma in humans; and furthermore, these models are valuable for evaluating surgical techniques and therapeutic drugs.29,30 For instance, the morphology of the wound and the efficacy of vitrectomy have been studied in this animal model of a contusion injury. Because of initial uveal engorgement and inflammatory swelling, early surgical intervention was hazardous. The findings support the clinical impression that vitrectomy in traumatized eyes with a substantial contusive component is best delayed for 1 or 2 weeks.26,27 (This is discussed in more detail in Chapter 110, Surgery for ocular trauma.)
A human RPE culture system is another useful means of investigating the RPE cell’s behavioral patterns, such as the migration, proliferation, and alteration of its phenotype, and the growth factors and cytokine-secreting patterns in order to understand the process of PVR.31–36
Wound healing and traumatic proliferative vitreoretinopathy
Wound healing in the eye is similar to that in other bodily tissues, consisting of three phases: exudation/inflammation, proliferation, and regeneration.37 The typical wound healing response in the anatomical setting of the eye and the vitreoretinal relationship explain the development of traction retinal detachment after penetrating ocular injury.
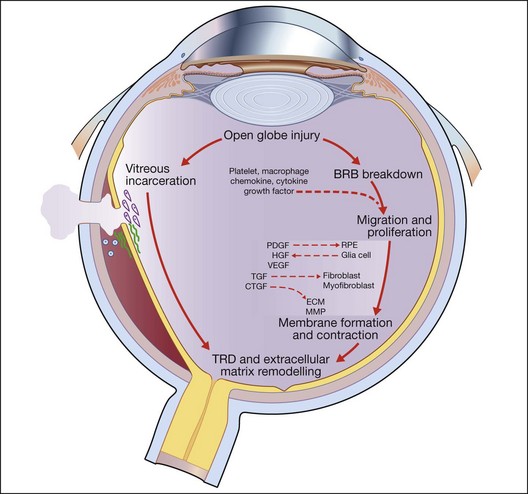
Open-globe injury results in a breakdown of the blood–retinal barrier and allows the entry of a variety of cells into the intraocular milieu, causing the expression of a variety of chemokines, inflammatory cytokines, and growth factors that affect the adjacent RPE, fibroblasts, and glial cells. In response, these previously resting cells undergo proliferation and migration as they change their pattern of gene expression, resulting in alterations of their own cytokine, extracellular matrix, and receptor profiles. Some cells – myofibroblasts, for example – proliferate and produce strong contractile forces that oppose the physiological forces that normally keep the retina attached and a tractional retinal detachment occurs. Following the natural course, proliferation is accompanied by a progressive accumulation of extracellular collagen and by a decrease in inflammation and inflammatory mediators.18,20 This wound healing process is central to the final common pathway that leads to traction retinal detachment and posttraumatic PVR in open globe injury (Fig. 98.3).
Cellular constituents
Epiretinal membranes removed during vitreous surgery for PVR after injury have been analyzed to gain an understanding of the origin and characteristics of their cellular constituents.38–40 Depending on the nature of the injury and the stage of the response, these membranes contain variable numbers of cells that are phenotypically identified as inflammatory cells, RPE cells, glial cells, fibroblasts, and myofibroblasts.
Inflammatory cells are among the earliest cell types to appear in the wound healing response. They may be attracted to the wound by chemokines upregulated in traumatized retinal tissue, by breakdown of the blood–retinal barrier, or as a response to intraocular blood.31,41–45 The cytokine products of these cells may be critical for the activation of other retinal cell types, further recruitment of inflammatory cells, and formation of collagen. Macrophages are a constant feature of experimental traction retinal detachment.42,43 In the primate model of posterior penetrating injury, macrophages are present before the invasion and proliferation of fibroblast or RPE cells.19,20 The finding of cellular and humoral immune responses to retinal antigens following retinal detachment and experimental PVR has suggested the possibility of an autoimmune component in PVR, although the evidence for a pathogenic role of such a response is incomplete.45–48
The RPE cell is central to the pathophysiological response seen in posttraumatic PVR. The RPE cells have critical characteristics, including migration and proliferation.32,33,38 The posttraumatic stimuli that are specifically responsible for RPE changes are not completely understood. RPE cell growth appears to be regulated by both paracrine and autocrine stimulation. RPE cells were shown to proliferate and form multilayered colonies of dedifferentiated RPE cells within 24 hours of retinal detachment in cats.49 They were also consistently found in the membranes of animal models of penetrating injury.20,22,26 Additionally, cultured RPE cells, just like RPE cells under the detached retina, produce mitogenic and chemotactic growth factors, such as platelet-derived growth factor (PDGF) and hepatocyte growth factor (HGF), and possess receptors for each of these growth factors. RPE cells respond not only to growth factors from RPE cells (autocrine) but also to those from surrounding tissue or from serum (paracrine), resulting in recruitment of additional RPE cells, and thus augmenting the process.34–36,50–56 In response to these stimulations, RPE cells may alter their phenotype to cells with a macrophage, fibroblast, or myofibroblast morphology.32,33,38 This fibroblastic RPE may synthesize and remodel the matrix on the retinal surface, contributing to the formation of the membrane. It was demonstrated that the proportion of RPE in human membranes varies according to the age of the membrane. The number of RPE cells is greater in early (<4 months) specimens and declines progressively as the membranes mature with more advanced extracellular matrices.57
Glial cells, identified by their typical morphologic characteristics and immunoreactivity to glial fibrillary acidic protein (GFAP), were found in neurosensory retina and membrane from full-thickness retinectomy specimens obtained at surgery for PVR, with increased expression correlated to the severity of degeneration after trauma.57–60 Glial cells appear to be involved in PVR formation through migration onto the surface of the retina and may be involved in remodeling of intraretinal synapses, possibly contributing to visual recovery after retinal injury.
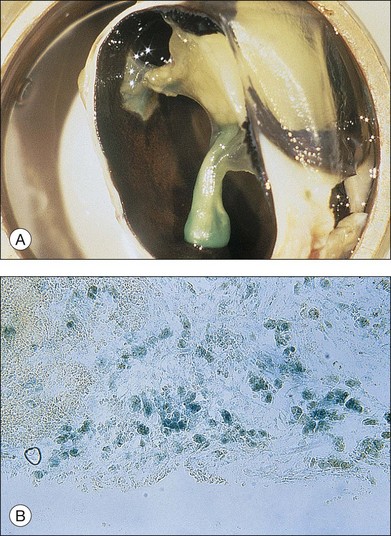
Fibroblastic proliferation is critical to the progression of post-traumatic proliferative response. Although fibroblasts are typically derived from RPE and glial cells, the immunohistochemical markers for these cells are missing in some fibroblasts, making their derivation uncertain.38,61,62 In studies using an animal model of globe perforation with long posterior wounds and injection of intravitreal blood, where membranes extended from the wounds and β-galactosidase labeled Tenon fibroblasts were identified in the vitreous and membrane, it was determined that at least some of the fibroblasts may have originated from Tenon’s layer at the wound edge (Fig. 98.4).20,24,25,63,64
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


