Chapter 71 The physiologic anatomy of eye muscles and the surgical anatomy of strabismus
The extraocular muscles
The six extraocular muscles (EOMs) form the effector arm of the ocular motor system which is a set of five distinct eye movement control systems (saccades, pursuits, vergences, and vestibulo-ocular and optokinetic reflexes). A common “motor plant” to all these different subsystems compriseds the EOMs, motor neurons, the globe, and the orbital connective tissue. The muscles are organized as “yoked pairs” that require precise coordination to maintain binocular fixation on visual targets. These muscles are highly specialized and functionally diverse with distinct biochemical, physiologic, and pathologic properties. The medial rectus (MR) and the lateral rectus (LR) muscles rotate the eye horizontally; adduction is accomplished by the MR and abduction by the LR. The two vertical rectus muscles and the two oblique muscles have more complex actions that depend on the direction of gaze and will be detailed later. The gaze position determines the effect of EOM contraction on the rotation of the globe. When the eye is in the primary position, more than 70% of the ocular motor neurons are active with an average firing rate of approximately 100 Hz;1 18% of motor neurons never cease firing for any eye position.2 This activity level is not common in other motor systems.
During embryogenesis, mesodermal primordia condense around the developing eye and proceed through the same myogenetic steps of other skeletal muscles. However, unlike skeletal muscles, the EOMs retain “developmental” protein isoforms, including embryonic myosin, and fetal acetylcholine receptor. The expression of these alternative proteins may underlie the susceptibility of the EOMs to disorders such as Graves’ disease and myasthenia gravis and their resistance to disorders such as muscular dystrophy. The EOMs have an enhanced antioxidant capacity and a substantial calcium sequestration capacity that protects them from toxic agents that act via increases in intracellular calcium concentrations, as well as other distinct functional and metabolic characteristics.3
The EOMs do not conform to traditional classifications of muscle fiber types which are based on the expression of myosin isoforms. EOMs are unique in that they possess different types of muscle cells. Each muscle cell is composed of groups of myofibrils called sarcomeres. A sarcomere is the major structural and functional unit of striated muscle. Actin and myosin arrangement results in a banding pattern seen by electron microscopy. Muscle contractions take place when myosin and actin filaments are actively moved past each other as a result of release of calcium by the sarcoplasmic reticulum. Fast twitch (“fibrillenstruktur”) muscle fibers generate fast eye movements and are composed of well-defined myofibrils with well-developed sarcomeres. “Felderstruktur” muscle fibrils generate slow or tonic eye movements and are composed of poorly developed sarcomeres. Cholinergic motor neurons supply both types of muscle fibers. The innervation of fibrillenstruktur fibrils is thick and heavily myelinated, with a single neuromuscular junction, whereas the innervation of felderstruktur fibrils is thin, with multiple grapelike clusters of neuromuscular junctions. The previous histologic classification of human EOMs has now been replaced by a more complex system based on global and orbital layer distribution, innervation status (single vs. multiple nerve contacts per fiber), and mitochondrial/oxidative enzyme content.3 Using this classification, five distinct EOM fiber types are identified:
1. Orbital singly innervated fibers (SIFs).
2. Orbital multiply innervated fibers (MIFs).
4. Global pale (granular) SIFs.
The predominant fiber in the orbital layer is the fast, twitching-generating, SIF type. These fibers are specialized for intense oxidative metabolism and fatigue resistance and are high in mitochondrial content. Twenty percent of the fibers are multiply innervated non-twitch fibers.4 It has been proposed that the role of these fibers is proprioception. Many individual muscle fibers do not extend the entire length of the muscle; only a few of the SIFs extend through the entire length of the muscle in the global layer. MIFs run the entire length of the muscle, even extending into the distal tendon. The orbital layer fibers are smaller and shorter than the global layer fibers. All fibers are active at all times and are recruited differentially during different ocular movements.5 MIFs are responsible for tonic activity; SIFs are responsible for generating most of the force needed to generate a saccade, but they also have roles in fixation and pursuit movements.
The functional heterogeneity of the EOMs is manifest in the myosin heavy chain phenotype of the EOMs. In contrast to skeletal muscles, embryonic myosin isoforms are retained throughout life in orbital layer fibers of the EOMs. In mature EOMs, the fibers of the orbital layer are heterogeneous in their expression of developmental myosin isoforms, with the highest levels in the proximal and distal regions of each fiber.6 The normal development of the orbital SIFs is dependent on environmental cues during the critical periods of vision development. Early visual stimulus has an important regulatory influence in the myosin expression patterns of the EOMs and shape EOM maturation.7,8
Because strabismus surgery uses millimetric grading, strabismus surgeons pay special attention to the exact location of the insertion sites of the EOMs and their relationship with respect to each other. Starting from the MR the progressive insertion distance with respect to the limbus of the rectus muscle tendons on the sclera has been depicted by a continuous curved line (the spiral of Tillaux). These distances are highly variable9–13 (Fig. 71.1) and knowledge of normal range rather than a fixed number is essential (Fig. 71.2) (Table 71.1).
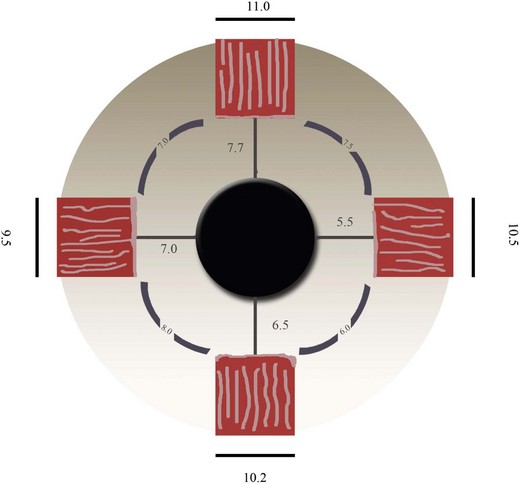
Helveston measured the insertion distance of the MR in 114 eyes and found a range between 3 and 6 mm, with an average of 4.4 mm.18 Kushner and Morton measured the distance from the limbus to the MR insertion in 80 eyes and found the range was between 3.5 and 5.5 mm, with an average of 4.3 mm.11 Keech measured 40 eyes of patients 10−30 months old with esotropia; the range was 5−6 mm (5.5 average). After disinserting the muscle, a movement of 1.2 mm on average toward the limbus was observed (0.5−2 mm).12
Axial length and age have been hypothesized as explanatory variables for anatomic variation and response to surgical dosages.19 Apt studied 100 cadaver eyes and failed to find a relationship between axial length and the EOM insertion distances. However, the age range was 21−90 years (average 60 years) therefore excluding children with immature structures and axial lengths less than 23 mm.9,15
The width of the muscle insertions (Table 71.1) is important when considering rectus muscle displacements and transpositions. “Full tendon-width” or “half tendon-width” are terms that are used for the surgical titration of these operations (usually 5 and 10 mm, respectively). However, full width transpositions are practically impossible without impinging on adjacent muscles.9
The EOMs are richly vascularized compared to skeletal muscles (Fig. 71.3). Two branches of the ophthalmic artery provide the blood supply to the EOMs: the lateral muscular branch and medial muscular branch. The LR, superior rectus (SR) and the superior oblique (SO) are supplied by the lateral muscular branch; the MR, inferior rectus (IR) and inferior oblique (IO) receive their blood supply from the medial muscular branch. The muscular branches give rise to the anterior ciliary arteries that pass from the muscle to the episclera and are responsible for the blood supply of the anterior segment of the eye. With the exception of the LR, all the EOMs have two ciliary arteries. Supplementary blood supply to the LR comes from the lacrimal artery and the infraorbital artery, which also supplies the IO and IR. When strabismus surgery involves several EOMs, the anterior segment circulation can be compromised. The venous system parallels the arterial system and empties in the superior and inferior orbital veins. The inferior and superior temporal vortex veins are important structures to be aware of during strabismus surgery. The inferior temporal vortex vein is located 8 mm posterior and temporal to the IR insertion. The superior temporal vortex vein is located near the posterior edge of the SO tendon insertion under the SR.
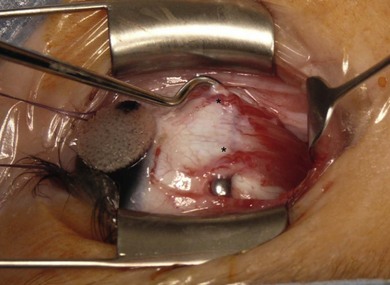
Fig. 71.3 Medial rectus muscle with two robust anterior ciliary arteries.
(Photo courtesy of Tina Rutar, MD.)



