Fig. 13.1
Schematic of cross-sectional view of a human lens showing various layers. The lens epithelium is a single layer of cells covering the anterior section. At the bow region, the epithelial cells elongate, lose their organelles, and differentiate to form fiber cells. The central nucleus is made up of primary fiber cells. The cortex comprises secondary fiber cells and is divided into inner and outer cortex. The outermost layer covering the entire lens is called the capsule
The lens derives all of its nutrients and oxygen from the aqueous humor and vitreous body. Because the lens is composed of fibers representing a range of ages, it is an attractive tissue for studying the effects of aging on protein structure and function [5]. Lens fiber cells are arranged in layers, with little extracellular space to disrupt the regular array [6]. Transparency is achieved by the tightly ordered packing and assembly of crystallins, the soluble proteins in lens fibers [7]. Protein concentrations in the lens are very high (450 mg/mL), yet there is no colloidal osmotic pressure (and hence no ability to draw in water). Both the packing and hydration properties of the proteins confer the refractive power to the lens. Although the lens has protective mechanisms that preserve its function during most of the human life span, these mechanisms begin to deteriorate with aging, leading to the gradual accumulation and aggregation of modified proteins, and a concomitant loss in the optical quality of the lens, and, ultimately, cataract formation [3, 8–11].
Crystallins make up about 90 % of the lens proteins and their properties account for the high refractive index of the lens [3, 7, 12]. Other proteins in the lens include cytoskeletal and membrane proteins, such as actin, filensin, and spectrin; transporters and channel proteins; proteins that constitute cell–cell junction and communication; and enzymes involved in metabolism, protein synthesis, and degradation [3, 7]. Crystallins produced within the lens during the formation of lens fiber cells have exceptional longevity. There are three classes of crystallins—α-, β-, and γ-crystallins—encoded by a diverse groups of genes. α-Crystallin constitutes 40 % of the total lens protein mass and is composed of two types of 20-kDa subunits, αA and αB. The αA- and αB-crystallins coexist, in roughly a 3:1 ratio, as polydisperse hetero-oligomers [12]. α-Crystallin belongs to the small heat shock protein (sHsp) family and functions as both a molecular chaperone and a structural protein [12, 13]. These properties are particularly relevant to the preservation of β- and γ-crystallins in the non-aggregating form to maintain lens transparency. β-Crystallins are a family of basic (βB1, βB2, βB3) and acidic (βA1, βA2, βA3, and βA4) polypeptides of 22–28 kDa. They exist as hetero-oligomers. γ-Crystallins exist only as monomers of 20 kDa. Both β- and γ-crystallins serve as structural proteins in lens [7].
13.2 Lens Proteases and Their Effect on Lens Proteins
Table 13.1 lists the various proteases and peptidases that have been identified or isolated from the lens and lens epithelial cells, underscoring the critical role the lens proteolytic system plays during lens development and morphogenesis [2, 4, 14, 15]. Most of the proteases expressed during lens development complete their major roles by the time the fiber cells attain maturity, but they remain in the lens, together with peptidases, and exert a low level of activity. With time, their cumulative effects contribute to the age-related truncation of crystallins, the cleavage of cytoskeletal proteins, membrane proteins such as aquaporins and connexins, and the generation of crystallin fragments. A number of proteases have been implicated in various aspects of lens development, organelle degradation, and lens physiology. Several endopeptidase activities have been demonstrated in mature lens fiber cells [16–25].
Table 13.1
Proteases and peptidases in lens and lens epithelial cells
Name | References |
|---|---|
Acylpeptide hydrolase | [26] |
ADAM-12, 19, ADAMTS7 | [27] |
Aminopeptidase III | |
Calpains | |
Caspases 3, 6, 7 | [35] |
Cathepsin B, D | [36] |
Dipeptidase | [37] |
Leucine aminopeptidase | [38] |
Matrix metalloproteases | |
Membrane proteinase | [21] |
Prolyl oligopeptidase | [41] |
Proteasome | |
Secretases | [46] |
Serine-type protease | |
Thimet oligopeptidase | [18] |
Trypsin-like protease |
To highlight some of the other evidence linking proteolytic activity to protein aggregation preceding cataractogenesis, proteases that cause truncation of α-crystallin missing five residues from its C-terminus have been identified as calpains [48]. In addition, in studies of cataract formation in rat lenses, calpains have been shown to induce accelerated truncation in α- and β-crystallins [49]. Proteolytic activity that cleaves αA-, αB-, and βA3/A1-crystallin to generate peptide fragments has been demonstrated in human lenses [19, 50]. Matrix-assisted laser desorption ionization imaging mass spectrometry (MALDI-MS imaging) studies have shown age-related changes in crystallin and other proteins in various species, including bovine, rabbit, guinea pig, rat and human lenses [51–55]. MALDI imaging studies have shown progressive truncation of αA- and αB-crystallins and aquaporin in concentric layers of fiber cell, with the highest cleavage in the oldest region of the lens [53, 55]. Further, the abundance of cleaved products increase with age in a given lens reinforcing the likelihood that an age-related increase in crystallin degradation occurs during aging [56]. The major bonds cleaved in vivo are on the carboxyl side of Asn101, Asp151, Ser168, and Ser169 of the A chain and on the carboxyl side of Thr170 of the B chain of bovine α-crystallin [11, 57]. The other truncation observed in αA-crystallin is due to cleavage at the carboxyl side of Ser172 [58], and the amount of cleaved αA- and αB-crystallin is high in the water-insoluble (WIS) fraction than in water-soluble (WS) fraction [59–61]. One study found that in aged human lenses, αA-crystallin is truncated by three residues and αB is truncated by one or six residues at the N-terminus [62]. Mass spectrometric analysis of WS and WIS fractions has identified several of the previously known sites of truncation as well as a few additional ones [60, 63–68]. But the protease(s) responsible for α-crystallin truncation at multiple sites has not been identified. Truncation of βB1-, βA3/A1-, and βB2-crystallins in vivo has been reported, based on the identification of several truncated and cleaved β- and γ-crystallins in both WS and WIS fractions of human lenses [59, 60]. Since truncation alters the stability of βB2-crystallin in vitro [49], it is hypothesized that in vivo βB2-crystallin truncation may be responsible for its aggregation with aging.
Proteasome is the main intracellular proteolytic system responsible for clearing the damaged proteins. Lens fiber cells and lens epithelial cells have a completely functional ubiquitin proteasome pathway (UPP), although the protein levels and activities of the protein components are lower in the inner region of the lens than in the outer region [42, 69]. The age-dependent decrease in proteasome activity in human lens may partly reflect a decreased rate of clearance of damaged proteins and peptides [70]. The ubiquitin-dependent proteolytic pathway in the lens is involved in the degradation of oxidatively modified proteins. However, due to an age-related decrease in ubiquitin conjugation activity, the oxidatively modified crystallins accumulate in the lens, primarily in the nuclear region. The accumulation of modified crystallins is believed to be responsible, at least in part, for the development of age-related protein aggregation and light scattering [9, 69]. Recent studies have shown that inhibition of the proteasome activity involved in the differentiation of lens fiber cells prevents normal maturation of fiber cells [15]. While calpains have been implicated in the maturation of rodent lens crystallins [49] and to some extent in the development of cataract in sheep [34, 63], the role of calpains in human cataractogenesis is not yet established because human lens also contains calpain inhibitor [30, 71]. The presence of lens proteins cleaved at specific sites during aging [57] provides credence to the idea that, although protease activity begins to decline in lens fibers as they mature, the aging lens nevertheless has appreciable levels of protease and peptidase activity [14, 29, 72]. Such activity may be exemplified in posterior capsular opacification (PCO) after cataract surgery. Proteases, such as matrix metalloproteases (MMPs), have been implicated in the development of PCO. In addition, the increased expression of TIMP3, ADAM-12, ADAM-19, and ADAMTS7 genes in response to transforming growth factor β (TGFβ) treatment suggests that these proteases might play a role in the development of PCO as well [27].
13.3 Effect of Proteolysis on α-Crystallin Subunits and Chaperone Activity
The lens WIS fraction (which is mostly composed of α-crystallin aggregates) contains greater amounts of C-terminally truncated αA- and αB-crystallins than does the WS fraction [58, 60–62, 67]. Additionally, C-terminally truncated α-crystallin has reduced chaperone activity [73, 74], suggesting that C-terminal truncation affects oligomerization of α-crystallin and its chaperone activity. Studies of the oligomeric size, structure, hydrodynamic properties, and chaperone function of the C-terminally truncated human αA-crystallin mutants suggest that truncation of crystallins plays a role in high-molecular-weight aggregate formation and loss of chaperone activity in the aging lens [73–75]. A portion of αA-crystallin is in its deamidated form in the aged lens. Surprisingly, a study of truncated αA-crystallin in its deamidated form revealed that the C-terminally truncated (αA-1-140) and deamidated (αA-N123D) form possesses better chaperone activity than the truncated form [76], raising the possibility that deamidation in a truncated αA-crystallin might be beneficial. In vitro studies have shown that the C-terminally truncated αA-crystallin (αA-1-162) is susceptible to degradation by the UPP. However, oligomerization of the truncated αA-crystallin with wild-type αA- or αB-crystallin was found to render it more resistant to UPP, perhaps one of the reasons for the accumulation of truncated crystallins in vivo [77].
The effect of α-crystallin truncation on oligomerization is apparent from our studies with recombinant αA151-crystallin. The addition of increasing amounts of αA151 to wild-type αA-crystallin causes a concentration-dependent increase in the oligomeric mass of αA-crystallin. When wild-type αA-crystallin and αA151 were mixed at a 3:1 ratio, the mass of the hetero-oligomers was 14.0 ± 0.04e5, whereas the oligomers formed when αA- and αA151 were mixed at 2:1 ratio was 20.0 ± 0.01e5 and both these masses were significantly higher than the mass of oligomers formed by wild-type αA-crystallin. There was also an increase in the hydrodynamic radius of αA-crystallin + αA151 oligomers (11.8 ± 0.3 nm), as compared to the hydrodynamic radius of oligomers of wild-type αA-crystallin (8.0 ± 0.2 nm). While the reasons for the increase in the size of oligomers in a dynamic α-crystallin molecule is not known, it might be due to the change in the subunit exchange rate following truncation. When labeled full-length αA + truncated αA151 hetero-oligomers, prepared at 2:1 ratio, were examined by fluorescence energy transfer assay, the subunit exchange rate was 0.0486/min, significantly lower than the 0.0572/min in the control experiment, suggesting that the C-terminal truncation affects the dynamic state of α-crystallin. Thus, the slower exchange rate may result in the retention of a larger number of subunits bound to the oligomer, leading to larger aggregate formation.
13.4 Analysis of Human Lenses and Identification of Crystallin Fragments
The concentration of crystallin fragments (<18 kDa) is higher in aged and cataract lenses than in normal young lenses [29, 49, 52, 59, 60, 66, 78, 79]. Similarly, we found that the concentration of low-molecular-weight crystallin fragments (<3 kDa) is greater in aged and cataractous lenses than in young lenses [56, 80]. In our studies of peptide levels in young lenses (20 years old; n = 5), aged lenses (70 years old; n = 5), and cataract lenses (74 years old; n = 5), the total peptide level in young lenses was 0.29 mg/lens, whereas in old and cataract (with light brown nucleus) lenses peptide levels were only marginally different (0.9 mg/lens and 1.1 mg/lens, respectively). A study of peptide distribution within the lens by MALDI tissue imaging showed that the inner most region of the lens, which represents the oldest lens fiber cells, has the greatest amount of peptides, suggesting an age-dependent accumulation of the peptides in vivo [9, 52, 65].
Our liquid chromatography–mass spectrometry (LC-MS) studies clearly point to age-dependent peptide levels in the lens, with aged lenses having a higher concentration than young and middle-aged lenses. The total ion current (TIC) elution profile of WS peptides in young, middle-aged, and aged lenses demonstrated more peptides in the aged lens than in the middle-aged lens. When the peptide ion populations from LC-MS data sets of lens samples from young and middle-aged lenses were analyzed by Agilent Mass Profiler software and the peptides showing statistically significant abundance changes across samples were plotted in a log/log plot, a total of 1,261 peptides was found in WS lens fractions and majority of them at a higher concentration in the WS fraction of middle-aged lenses. When the data were filtered to exclude peaks whose abundance was <1 % of the most abundant, there were 128 peptides, of which 18 were unique to young lenses and 102 were unique to middle-aged lenses, and only 8 peptides were shared (Fig. 13.2a). Thus, there was a fivefold increase in the number of distinct peptides in middle-aged lenses when compared to young lenses. In the comparison between middle-aged and aged lenses, 1,804 peptides were detected and they were clearly more abundant in aged lenses than middle-aged lenses. Further analysis showed that 78 peptides were unique to middle-aged lenses, 199 unique to aged lenses, and only 74 peptides were shared (Fig. 13.2b). Thus, there is a clear progression of peptide appearance from young to aged lenses. Interesting results also emerged from our LC-MS analyses of peptides in the urea-soluble fraction of the lens proteins from different age groups. In the young vs. middle-aged comparison of peptides, 718 peptides were detected. When data were filtered to exclude peaks whose abundance is <1 % of the most abundant, only 79 peptides differentially abundant in both samples were detected and most of the peptides appear in middle-aged lenses (Fig. 13.2c). This finding indicates the prominent presence of WIS, urea-soluble peptides in middle-aged lenses. In the middle-aged vs. aged comparison, 1,502 peptides were detected with a bias towards middle-aged lens. However, after applying the 1 % threshold to the data, there were 195 peptides, of which 13 were unique to middle-aged lenses, 157 unique to aged lenses, and 25 were shared (Fig. 13.2d). Thus, the age-related increase in peptide abundance in both WS and urea-soluble fractions of the lens attests to an age-dependent increase in the cleavage of crystallins.
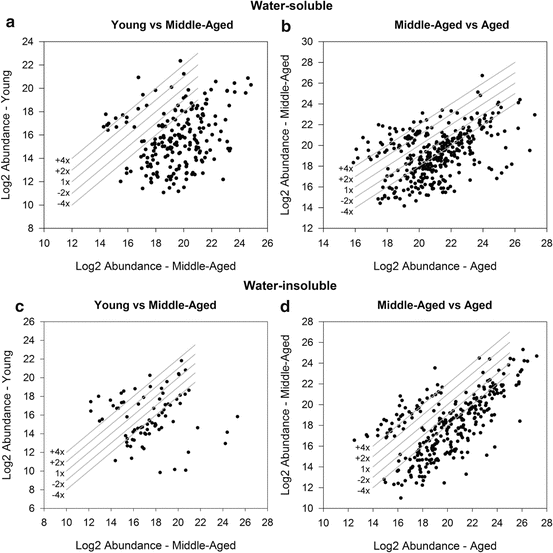
Fig. 13.2
A log/log plot displaying fold changes in water-soluble (WS) (a, b) and water-insoluble (WIS) (c, d) peptides in young vs. middle-aged lenses (a, c) and middle-aged vs. aged human lenses (b, d). The WS and WIS fractions from the lenses were treated with 6 M urea and filtered using 10 kDa filter to isolate the LMW peptides. The peptides were analyzed by LC-MS on an Agilent 6520 Accurate Mass QTOF mass spectrometer at University of Missouri Proteomic Core facility
A number of factors could be involved in the accumulation of low-molecular-weight peptides (<10 kDa) with age, such as the activation of proteases, decreased clearing of the peptides due to diminished peptidase activities, the binding of the peptides to crystallins, and making them resistant to further degradation by peptidases, and increased nonenzymatic cleavage of crystallins in aging lenses due to free radicals generated in vivo. The presence of latent protease and nonenzymatic cleavage of lens crystallins is well documented [20, 64, 68]. Our studies with lens acylpeptide hydrolase (APH) demonstrate that a truncated form of this enzyme has the potential to cleave lens proteins and it is present in aged lenses [26, 81, 82]. Analysis of different regions of the bovine lens revealed the least amount of peptidase activity in older lens fiber cells with the highest amount of crystallin fragments (Fig. 13.3). Further, on the basis of the activity distribution of major peptidases in the lens, it is evident that the activity of leucine aminopeptidease, aminopeptidase III, prolyloligopeptidase, and APH, whose action together would result in complete hydrolysis of any peptide, is significantly less in the nuclear region as compared to the outer and inner cortical region of the same lens (Fig. 13.3).
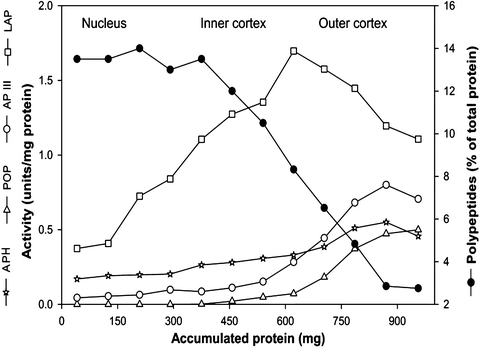
Fig. 13.3
Relationship between lens polypeptide levels and protease activities in different regions of the bovine lens. Decapsulated lenses were agitated in 3 mL portions of water and, after certain amount of fiber cells were dissolved, the undissolved portion of the lens was transferred to another container and the process was repeated 12 times to completely dissolve the lens. The 12 fractions were further homogenized in a dounce homogenizer. Leucine aminopeptidase (LAP) activity was assayed at pH 8.5 using Leu-pNA after activation with MgCl2. Aminopeptidase III (APIII), acylpeptide hydrolase (APH), and prolyl oligopeptidase (POP) activities were assayed at pH 7.5 using Arg-pNA, N-Acetyl-Ala-pNA, and tBOC-Ala-Ala-pNA as substrates, respectively. The polypeptide (<18 kDa) content in each fraction was measured following SDS-PAGE by image analysis. The polypeptide levels decrease with increasing protease activities and vice versa suggesting a correlation between accumulation of lens polypeptide and proteolytic activity of lens
We have identified a large number of the peptides in human lenses of different age groups [56, 80]. Several of the peptides were from the N-terminal domain of αA- or αB-crystallins or the chaperone site [56, 83]. There were also representative peptides from β- and γ-crystallins. However, it has not been possible from the peptides identified to reconstruct the full sequence of crystallins, suggesting that some of the peptides are fully hydrolyzed into amino acids. How all of these peptides are formed and why some are not completely hydrolyzed into amino acids in vivo are not known. Specific proteases are likely involved in the generation of some peptides, whereas other peptides are likely generated by free radical-mediated cleavage of peptide bonds. Recently, we synthesized peptide substrates with αA-62-70 and αA-76-85 sequence and demonstrated that the lens has proteases that cleave these synthesized substrates at the site(s) corresponding to the in vivo cleavage that results in the release of αA-66-80 peptide from αA-crystallin [25]. Human lens has αA-66-80 and βA3-188-215 peptides [56] and their truncated forms, likely generated by the action of aminopeptidases. A separate study using synthetic peptides showed that spontaneous cleavage of peptide bonds might be occurring to generate peptides with Ser-terminal residues [64].
13.5 Crystallin-Derived Peptides Induce Protein Aggregation
Proteolysis has been implicated as one of the cataract-inducing events [14, 84]. However, until recently, the molecular mechanism of crystallin fragment-induced protein aggregation in the lens has not been explained, although the intrinsic aggregation propensity of amyloidogenic peptides has been shown to correlate with protein aggregation disease [85, 86]. We were the first to demonstrate that peptides derived from oxidized crystallins induce the aggregation of crystallins by interaction at specific regions [87]. Subsequently we found that αB-1-18 (MDIAIHHPWIRRPFFPFH) and βA3/A1-102-117 (SD(N)AYHIERLMSFRPIC) peptides, present in aged and cataract lenses, induce aggregation of αB-crystallin [87, 88]. The αB-crystallin complexed with βA3/A1-102-117 peptide showed diminished chaperone activity, thereby preventing its ability to suppress protein aggregation. Peptide cross-linking and mass spectrometric studies showed that the βA3/A1 102-117 peptide interacted with αB-crystallin regions 70-74, 83-107, 121-123, 150-157, and 164-175, which are part of the α-crystallin domain and C-terminal extension. These data suggest that the βA3/A1-102-117 peptide acts like an anti-chaperone peptide because of its ability to interact with αB-crystallin at the chaperone site [88].
Our extensive studies with αA66-80 peptide and its truncated forms αA-66-75, αA-67-75, and αA-67-80, which are present in aged lenses and in increasing amounts in cataract lenses [56, 80, 83] show that these peptides interact with purified α-crystallin and cause aggregation and precipitation (Fig. 13.4). Of all the crystallin-derived peptides, αA-66-80 was found to be the most effective in precipitating α-crystallin (~70 %). Among the three truncated αA-66-80 peptides, αA-67-80 precipitated and aggregated the α-crystallin the most (49 %), followed by αA-67-75 (30 %) and αA-66-75 (20 %). Under similar experimental conditions, a peptide from a different region of αA-crystallin (αA-43-56) caused only 10 % protein precipitation. The interaction of the αA-66-80 peptide with α-crystallin also diminished α-crystallin chaperone activity pointing to one of the peptide interaction site(s) in α-crystallin subunits as a chaperone site, which was indeed subsequently confirmed by isotope-labeled cross-linking and mass spectroscopic studies with αB-crystallin and αA-66-80 [83]. The αB-70-107 and 164-174 regions were identified as αA-66-80 binding sites, and αB-72-92 region is now known to be one of the chaperone sites in αB-crystallin [89].
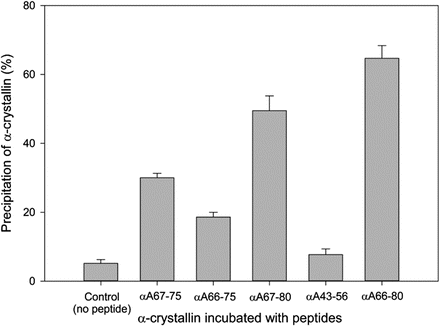
Fig. 13.4
Certain crystallin peptides found in the aged human lens precipitate α-crystallin in vitro. α-Crystallin and peptides in the ratio of 2:1 (w/w) were incubated in 500 μL of 50 mM phosphate buffer (in duplicates) at 37 °C for 24 h. After incubation, samples are centrifuged at 8,000 rpm for 30 min. Pellets were re-dissolved in 20 μL of 6 M urea and made up to 200 μL with buffer. Protein concentration of the re-dissolved pellets was estimated by Bio-Rad protein assay reagent
13.6 Mechanism of Proteolysis-Mediated Crystallin Aggregation and Development of Cataract
Data from in vitro and in vivo experiments show that lens opacity resulting from crystallin protein aggregation bears the hallmarks of increased proteolysis and decreased α-crystallin chaperone activity, features observed in aging cataract-prone lenses [9, 11, 33–35, 49, 55, 60, 65, 73, 74]. Proteins from aging lenses show increased posttranslational modifications. As depicted in the Fig. 13.5, it can be argued that the crystallin fragments accumulating in vivo due to proteolysis (enzymatic + nonenzymatic) occupy the central position in the pathway for lens opacification because of both their ability to induce aggregation and precipitation of α-crystallin and their anti-chaperone activity [56, 80, 83, 87, 88, 90]. The interaction of crystallin-derived peptides with α-crystallin can bring about structural changes that encompass hydrophobicity enhancement and the propensity to aggregation with additional crystallin molecules. These mechanisms are supported by our observation that after peptide interaction with α-crystallin, there is increased binding of hydrophobic site-specific probe bis-ANS [83]. Further, time-lapse recording has shown that α-crystallin–peptide complexes, once formed, attract additional protein molecules and the aggregates begin to grow [80]. The importance of the interactions between α-crystallin and crystallin-derived peptides and the accumulation of crystallin fragments as a result of proteolysis garners additional support from the observation of higher amounts of crystallin fragments in transgenic lenses expressing the protease APH prior to the development of lens opacity [91].
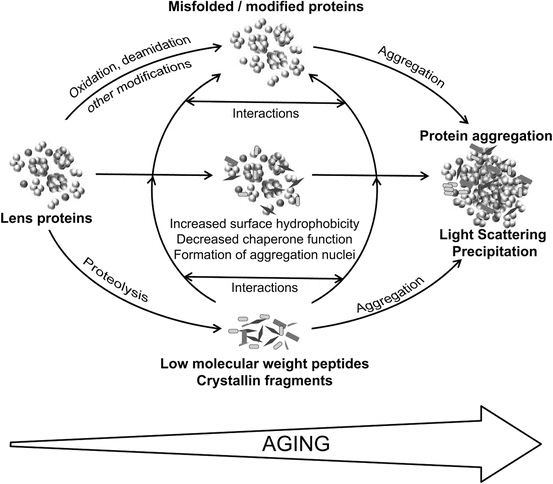
Fig. 13.5




Schematic representation of the proposed role of proteases and crystallin fragments in lens aging and cataract formation
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
