CHAPTER 151 Noise-Induced Hearing Loss
One of the most common causes of permanent hearing impairment is exposure to excessive sounds. Millions of individuals worldwide have noise-induced hearing loss (NIHL), resulting in a reduced quality of life because of social isolation, and possible inexorable tinnitus and impaired communication with family members, coworkers, and friends.1 The costs in terms of compensation and early retirement payments for work-related NIHL are immense. The U.S. Department of Veterans Affairs spends approximately $700 million a year on disability compensation and treatments for NIHL. NIHL is the single largest disability expenditure of the Veterans Benefits Administration.2
Measurement of Noise
The term noise is commonly used to designate an undesirable sound. In the scientific and clinical fields that deal with hearing, this term has come to mean any excessively loud sound that has the potential to harm hearing. The temporal patterns of environmental noise are typically described as continuous, fluctuating, intermittent, or impulsive.3 Continuous or steady-state noise remains relatively constant, whereas fluctuating noise increases and decreases in level over time, and intermittent sounds are interrupted for varying time periods. Impulsive or impact noises caused by explosive or metal-on-metal mechanical events have rapidly changing pressure characteristics consisting of intense, short-lasting (i.e., milliseconds) wave fronts, followed by much smaller reverberations and echoes that occur over many seconds. The amount of noise, usually referred to as the sound pressure level (SPL), is conventionally measured by a sound-level meter in decibel (dB) units using a frequency-weighting formula called the A-scale. The dBA-scale metric of sound level essentially mimics the threshold-sensitivity curve for the human ear, so the low-frequency and high-frequency components are given less emphasis as auditory hazards. Standard sound-level meters have electronic networks designed to measure noise magnitude automatically in dBA, whereas to measure impulse or impact noise, a more intricate peak-reading sound-level meter is needed that is capable of accurately measuring sounds with essentially instantaneous onset times.
Nature of the Hearing Loss
In PTS, the elevation in hearing thresholds is irreversible because lasting structural damage occurs to the critical elements of the cochlea. The precise relationship between the TTS and PTS stages of hearing loss caused by noise exposure is unknown. Although it seems logical to assume that repeated episodes of TTS would eventually lead to PTS, experimental findings imply that the fundamental processes underlying the development of reversible versus permanent NIHL are unrelated. Nordmann and colleagues4 using a survival fixation approach showed that the histopathologic manifestations of TTS and PTS noise damage to the chinchilla cochlea are distinct. Specifically, TTS was correlated with a buckling of the supporting pillar cell bodies in the frequency region of the maximal exposure effect. The morphologic abnormality that was consistently correlated with PTS was a focal loss of hair cells, and a complete degeneration of the corresponding population of nerve fiber endings. Because PTS eventually develops from repeated exposures to stimuli that initially produce only TTS, it is likely that the latter condition is also associated with subtle changes to the sensitive outer hair cell (OHC) system that go undetected by conventional light microscopy.
Acoustic trauma was previously a relatively rare event that was typically associated with accidental explosions in industrial settings. Military servicemen and servicewomen caught in roadside bomb explosions in the current armed conflicts in Iraq and Afghanistan are returning home in epidemic numbers, however, with profound permanent hearing losses and tinnitus.5 Consequently, acoustic trauma is a hearing problem that is increasing, at least in combat troops. Because many of these postdeployment cases are being treated in the private sector, all otolaryngologists may see acoustic trauma in increasing numbers.
Irreversible NIHL is a specific pathologic state exhibiting a recognized set of symptoms and objective findings.6 NIHL includes (1) a permanent sensorineural hearing loss with damage principally to cochlear hair cells, and primarily to OHCs; (2) a history of a long-term exposure to dangerous noise levels (i.e., >90 dBA for 8 hours/day) sufficient to cause the degree and pattern of hearing loss described by audiologic findings; (3) a gradual loss of hearing over the first 5 to 10 years of exposure; (4) a hearing loss involving initially the higher frequencies from 3 to 8 kHz before including frequencies less than or equal to 2 kHz; (5) speech-recognition scores that are consistent with the audiometric loss; and (6) a hearing loss that stabilizes after the noise exposure is terminated.
A patient with NIHL commonly consults a physician because of difficulties in hearing and understanding ordinary speech, especially in the presence of background noise. Many variations can be found in the detailed configuration of the audiogram of a noise-damaged ear, depending on the temporal and spectral distribution of the noise stimulus, and on the stage of hearing loss. The pattern of hearing loss most commonly associated with the earlier stages of NIHL is illustrated in Figure 151-1A. The beginning region of impairment involves the sensitive midfrequency range, primarily 3 to 6 kHz, and the corresponding hearing loss is classically described as the “4-kHz notch.” This pattern of maximal hearing loss, with little or no loss at less than 2 kHz, typically occurs regardless of the noise-exposure environment. The audiogram results in Figure 151-1A also show the sensorineural aspect of NIHL in that thresholds for bone-conducted stimuli are essentially identical to the thresholds for air conduction. The profile of noise-induced threshold hearing is usually symmetric for both ears, particularly for individuals who have been working in noisy industrial settings in which there are “surround” sounds.
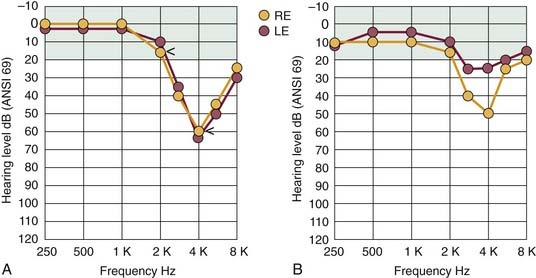
Commonly, other forms of noxious sound, such as the gunfire associated with sport shooting, cause an asymmetric pattern of hearing loss similar to the one illustrated in Figure 151-1B. In this case, the ear pointed toward the source of noise (gun barrel), which is the right ear of the left-handed shooter depicted in Figure 151-1B, would have worse hearing than the ear directed away from the source (in this example, the left or protected ear) by 15 to 30 dB or more, and particularly at higher frequencies because of the absence of the protective head-shadow effect.
The development of a hearing loss caused by habitual exposure to moderately intense levels of noise typically consists of two stages. Initially, the middle to high frequencies exhibit the resulting hearing loss. As the length of time of exposure to loud noise increases, hearing loss becomes greater and begins to affect adjacent higher and lower frequencies. In a classic cross-sectional study of occupational NIHL, Taylor and colleagues7 showed the gradual loss of hearing sensitivity in workers caused by habitual exposure to the intense sounds of dropforging tools that are commonly used in foundries. Figure 151-2 illustrates the progressive effects of exposure to the wideband noise (Fig. 151-2A) of two types of forging tools, presses and hammers, on the magnitude of hearing loss as the length of exposure increased. For the operators of the press and hammer equipment (Fig. 151-2B and C), the approximate 10- to 20-dB threshold shifts, typically observed at the higher frequencies during the first 1 to 2 years of exposure, grew to be a 20-dB or greater loss, from 3 to 6 kHz, after a 3-year exposure. There was a 40-dB or greater threshold shift after an 8-year exposure.
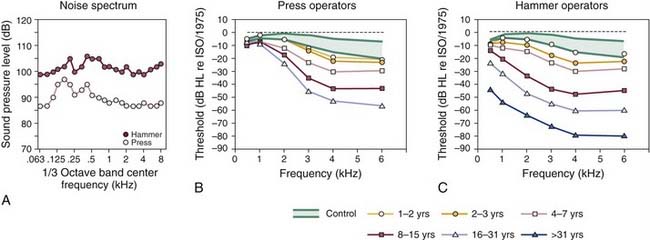
Detailed comparisons of the NIHL growth curves of Figure 151-2B and C reveal that with continuing exposure, hearing loss worsened at the higher frequencies and spread to the lower frequencies. In addition, for average exposure times of less than 10 years, hearing levels for the press and hammer operators who were exposed to mean levels of 108 and 99 dB SPL deteriorated similarly. For long-term exposures of 10 years or more, the results of Taylor and colleagues7 indicated that hearing losses resulting from the hammer-induced impact noise were greater than the losses resulting from the more continuous noise of the press equipment. Finally, a characteristic feature of NIHL, which is clearly documented in Figure 151-2B and C, is that hearing levels are increased rarely beyond about 70 to 90 dB of hearing loss, on average, even after more than 31 years of continuous noise exposure.
Cochlear Damage
Johnsson and Hawkins8 were among the first investigators to describe the typical patterns of cochlear injury for humans with chronic exposure to different types of loud sound. The photomicrograph of the cochlear tissue in Figure 151-3A depicts some common histopathologic consequences of NIHL for a patient with a lengthy history of noise exposure. The authors reported that the 50-year-old patient had worked intermittently over a 5- to 6-year period in an automotive body–stamping plant, and that he had a long history of using recreational firearms. The sharp transition in the basal end (following the uncoiling to the right) from the normal-looking organ of Corti (a darkish stripe corresponding to the region of inner hair cells and OHCs), with its dense network of nerve fibers, to the complete absence of hair cells and their corresponding nerve fibers (the much lighter adjacent area) can be noted. Figure 151-3B graphically reconstructs the histopathologic features of this cochlea as a cytocochleogram by depicting the number of remaining hair cells, in the form of percentages, averaged over 1-mm sections. A typical finding in individuals exposed to the occupational noise exemplified in this case is the almost symmetric pattern of degeneration observed for the two ears. The inset at the top right of the cytocochleogram shows the patient’s audiogram obtained about 1 year before his death, which shows the severity of the anatomic damage in functional terms by revealing an abrupt hearing loss for test frequencies greater than 2 kHz.
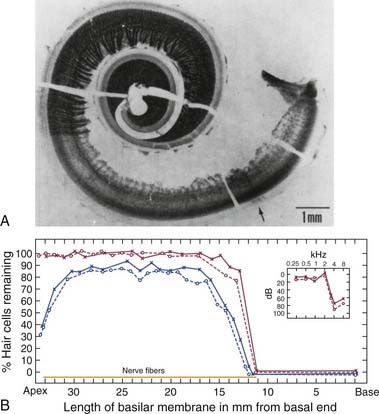
Examination of human temporal bone specimens by numerous laboratories8–10 has yielded documentation of the progressive stages of noise damage as depicted by epidemiologic data such as those of Figure 151-2. First, in the predictable sequence of events, a small region of hair cell and nerve fiber degeneration appears bilaterally at a cochlear region corresponding to the 4-kHz notch. Typically, these discrete lesions gradually grow in the basal-ward direction (i.e., toward the high-frequency extent of the cochlea) to involve a greater portion of the organ of Corti. Finally, as exposure to noise continues over years, the remaining sensory and neural elements in the basal end of the cochlea are destroyed, resulting in an abrupt loss of midfrequency to high-frequency hearing, such as that depicted by the clinical audiogram of Figure 151-3.
Research on Noise-Induced Hearing Loss
Scientific interest in the damaging effects of excessive sound on hearing has a long history for many reasons. First, the experimental strategy of exposing animals to noise and examining their ears for the sites of the resulting acoustic injury was used in the past as the independent variable in establishing some of our basic knowledge about hearing. Particularly, with the use of intense tones as the damaging agents, frequency information relating physical distance along the basilar membrane to the “best” frequency of the injured region provided an initial basis for understanding the tonotopicity of the cochlea and the central projection of such frequency-related information.11 Additionally, noise-damage strategies have been used to contribute to our understanding of the function of inner hair cells and OHCs by permitting differences in their central terminations in the ventral and dorsal cochlear nuclei to be distinguished.12
Anatomic Mechanisms Underlying Noise Damage
Experimental studies have led to an increased understanding of some major features of NIHL. It is well-accepted that the origin of the 4-kHz notch in the NIHL audiogram is related to the resonator function of the external auditory ear canal,13 rather than to indeterminable innate properties of the cochlea, such as a reduced vascular supply to this region of the organ of Corti.14 The primary research interest has always been, however, in the fundamental mechanism by which the sensory cell degenerates or is damaged after exposure. Numerous mechanisms15 have been proposed, including mechanical injury caused by severe motion of the basilar membrane, metabolic exhaustion of activated cells, activity-induced vascular narrowing that causes ischemia, and ionic poisoning from interruption of the normal chemical gradients of the cochlea owing to minuscule disruptions in the organization of sensory and supporting cells.
Although the many years of experimental research have not yet produced a major breakthrough in understanding of damage mechanisms, the current most convincing morphologic evidence supports a combination of the mechanochemical theories. First, at the ultrastructural level, it is likely that alterations in the stereocilia in the form of shortened or broken rootlets are involved in the initial pathologic processes that lead to TTS and, if such injuries are not repaired, then PTS.16–18 More recent findings showed that hair bundles are capable of rebuilding their ultrastructure from top to bottom over a 48-hour period.19,20 If damage is so severe that it overwhelms this self-repair mechanism as exposure continues, a discrete but direct mechanical disruption likely results in a toxic mixing of endolymph and perilymph through microbreaks in the structural framework of the cochlear duct,21 which leads to secondary effects, including loss of hair cells and their corresponding nerve fibers.
New Knowledge about Cellular and Molecular Mechanisms of Noise-Induced Hearing Loss
Hair Cell Regeneration and Repair
In the late 1980s, several seminal reports on hair cell regeneration in avian species established that hair cells in neonatal and adult birds regenerate after exposure to either damaging levels of sound22–24 or ototoxic antibiotics.25 Additionally, follow-up studies showed that recovery of cochlear function accompanied the cellular-recovery process.26–28 In the best-studied model for hair cell regeneration, the neonatal chick, it was shown that new hair cells arose as progeny from an otherwise nondividing supporting-cell population that was induced to proliferate by the damaging insult.29
Generally, noise-induced hair cell loss in the mammalian cochlea is irreversible. Experimental findings using in vitro cultures of neonatal mouse cochleas showed, however, that overexpression of mammalian atonal homolog 1 (Atoh1), also known as Math 1, a basic helix-loop-helix transcription factor known to be necessary for hair cell differentiation during development, leads to an increase in the production of extranumerary hair cells.30 Consequently, it seems that certain cells in the mammalian organ of Corti, at least in young animals, can be redirected toward a hair cell fate by the overexpression of Math 1.31 In addition, a most exciting finding was the discovery that new hair cells can be grown in a mature mammalian ear using the Math 1 gene. Kawamoto and colleagues32 injected an adenovirus carrying the Math 1 gene into the endolymph of mature guinea pigs. Using scanning electron microscopy, 1 to 2 months after the inoculation of the Math 1 gene insert, immature hair cells within the nonsensory supporting cells of the organ of Corti were detected. Most importantly, follow-up studies in the deafened guinea pig showed that not only did Math 1 induce regeneration of hair cells in the adult, deafened guinea pig, but also substantially improved hearing thresholds were measured.33 These findings that Math 1 (Atoh1) can direct hair cell differentiation and induce hearing function in mature nonsensory cells support the notion that adenoviral gene therapy based on expressing crucial developmental genes for cellular and functional restoration in damaged auditory epithelium may lead some day to a new treatment for NIHL.
Protection from Conditioning the Cochlear Efferent System
The outcomes of other experiments indicate that the mammalian cochlea may be capable of actively adapting to certain high-level sounds by receiving “exposure experience.” The notion that the cochlea can become resistant over time to the consequences of excessive sound was reported initially by noting “conditioning” effects in several animal models.34–37 The typical conditioning paradigm consists of providing a pre-exposure training experience using a moderate-level stimulus that, at a more intense level, becomes the subsequent overexposure stimulus. Together, these findings in animal models suggested that the mammalian cochlea might be capable under certain conditions of dynamically adapting to excessive sound.
For humans, the practical implications of the capacity to develop a “resistance” to loud sounds are obvious. A follow-up study in teenagers used a TTS-type paradigm to show the relevance of resistance training to humans. In this experiment,38 the investigators provided a pre-exposure training period in which the young subjects were exposed to 6 hours of pop/rock music at around 70 dBA. Threshold shifts in response to a 10-minute exposure to a 105-dB SPL, one-third octave-band noise centered at 1 kHz were compared for pretraining versus post-training intervals. The major result was that the “trained” ears exhibited significant decreases in TTS compared with their baseline values, showing that the “conditioning” effect, or the development of “resistance,” can be shown in human subjects, at least under brief TTS-exposure conditions.
Such human studies of the relationship between noise exposure and permanent hearing loss are more difficult to perform for many reasons including the ethical issues involved in the deliberate exposure of study subjects to noises, even though they are thought to be reversibly damaging. More recent research into the protective role of cochlear efferents in NIHL has depended more on the benefits of diagnostic testing with evoked otoacoustic emissions (OAEs) to predict potential susceptibility to the aftereffects of noise exposure.39 This simple, noninvasive, and objective procedure is based on the systematic measurement of a class of cochlear responses (i.e., the OAEs) that are primarily generated by the OHCs.40 Not only are OHCs exquisitely sensitive to the initial effects of acoustic overstimulation, which makes them excellent indicators of sound-induced ultrastructural damage, but also the final common pathway of the descending auditory-efferent system preferentially innervates the OHCs.
To take advantage of the ability of OAEs to measure efferent activity, several experimental paradigms were developed including a common one41 that uses contralateral acoustic stimulation to elicit medial olivocochlear-induced reductions of OAEs from the ipsilateral test ear. The other procedure42 is based on measurements of a subtype of evoked emissions represented by distortion product OAEs (DPOAEs) at 2f1-f2. In this strategy, long-lasting f1 and f2 primary tones of approximately 1 second are applied binaurally to elicit an efferent-based fast adaptation response that tests the ability of the combined medial and lateral cochlear-efferent systems to suppress DPOAEs in the test ear.
Experiments in guinea pigs documented the ability of the fast-adaptive DPOAE response to predict vulnerability to acoustic injury based on the vigor of efferent activity.43 The robustness of the olivocochlear efferents was inversely correlated with the degree of cochlear dysfunction after subsequent noise exposure, in that animals exhibiting large adaptive effects showed smaller postexposure losses than animals displaying small amounts of efferent-induced adaptation. It is clear from these findings that a modification of this assay could easily be applied to human populations to screen for individuals most at risk in noisy environments.
Some studies44 simply relating the presence or absence of the contralateral acoustic stimulation–induced reduction of emissions in human ears have reported such suppression is absent in most industrial workers who otherwise have normal OAEs. One implication of such observations is that a lack of efferent-related activity may be an early indication of cochlear damage from exposure to noise. Experimental studies such as these will eventually determine whether inherent efferent processes can account for and predict the remarkable individual variation in susceptibility to NIHL. The use of efferent-induced suppression of evoked OAEs in predicting susceptibility to NIHL, or in monitoring of noise effects in hearing-conservation programs, represents a promising line of future investigation.
Pharmacologic Protection from Noise-Induced Hearing Loss
It has long been recognized that hypoxia is a major pathogenic factor in NIHL. Based on the assumption that oxidative stress plays a substantial role in the genesis of noise-induced cochlear injuries that lead to permanent hearing loss, numerous pharmacologic strategies have been developed primarily in animal models to enhance the intrinsic defense mechanisms of the cochlea against this condition.45 Kopke and colleagues46 postulated several causes of noise-induced oxidative stress, all of which are amenable to pharmacologic treatments. Specifically, these investigators proposed that noise-induced oxidative stress leading to cochlear injury was related to (1) impaired mitochondrial function with respect to bioenergetics and biogenesis; (2) glutamate-induced excitotoxicity, which is the main excitatory neurotransmitter in the peripheral and central auditory systems; and (3) depletion of glutathione (GSH), an antioxidant that protects cells from toxins such as free radicals.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


