Fig. 8.1
Carcinine is efficiently transported to the retina following topical administration. Mice received carcinine dissolved in eye drop formulation (glycerol 1 %, carboxymethylcellulose 0.3 %, benzyl alcohol 0.3 %, in water, pH adjusted to 6.8 with potassium borate and potassium bicarbonate) [25]. (a) 0.2 M carcinine was administered at 0 min (arrow) and mice were euthanized 30, 60, 90, and 120 min later to collect retinas. Another set of mice received 0.2 M carcinine twice, at 0 and 60 min (indicated by arrows) and were euthanized every 30 min following second administration to collect retinas. (b) Mice received indicated concentrations of carcinine in eye drop formulation and were euthanized 30 min later to collect retinas. HPLC/MS was used to quantify the amount of carcinine per retina in each retinal sample [8]. The means of three retinal samples and error bars representing the standard error from the mean (SEM) were plotted
8.2.2 Carcinine Lowers 4-HNE-Protein Adduct in Retinal Cells
We previously demonstrated the scavenging effect of carcinine on 4-HNE in vitro [8]. To determine if carcinine had this effect in vivo, we measured the amount of 4-HNE-protein adduct in mouse retinas, following administrations of 0.1 M carcinine eye drops, every hour for 6 h [9]. After treatment with carcinine eye drops, a significant (P < 0.05) 20 % decrease of 4-HNE-protein adduct was found in mouse retinas when compared with vehicle-treated eyes [9]. Low amount of 4-HNE is constantly generated in cells and reacts with surrounding proteins to form 4-HNE-protein adduct within the intracellular compartment. Thus, our result supports the idea that carcinine is still active after migrating through the eye globe to the retina, and suggests that carcinine is entering the retinal cells, perhaps through a transport mechanism, where it prevents and/or reverses adduct formation.
8.2.3 Retinal Bioavailability of Carcinine
As shown in Fig. 8.1b, after application of 1 eye drop (10 μL) of 0.2 M carcinine corresponding to 2,000 nmol of carcinine on the mouse cornea, 100 ng of carcinine is found in the retina corresponding to 0.5 nmol. Thus, 0.025 % of the topically applied carcinine reaches the retina, which is at least 60-fold higher than the 0–0.0004 % intraocular bioavailability of most compounds [2]. In addition, topical applications of carcinine lead to therapeutic concentrations of carcinine, as demonstrated by the significant decrease of 4-HNE-protein adduct in the retina [9]. This result is interesting because if identified, the transport route allowing carcinine to efficiently penetrate the eye and reach the retina could be exploited for the delivery of other drugs to the posterior segment of the eye. Drugs can be transported through the corneal epithelium or through the conjunctival epithelium [2]. We used the HCECs to investigate the uptake of carcinine with the goal of identifying the transporter(s) involved.
8.3 Carcinine in HCECs
8.3.1 Carcinine Uptake in HCECs
We first hypothesized that the peptide transporters PepT1 (encoded by the SLC15A1 gene) and PepT2 (encoded by the SLC15A2 gene) could be involved in the transport of carcinine through the corneal epithelium because expression of SLC15A1 and SLC15A2 genes were detected in human cornea and HCECs [10, 11]. In addition, a recent study demonstrated that carnosine and anserine, two dipeptide derivatives structurally similar to carcinine, are transported with high affinity by PEPT2 and with moderate affinity by PEPT1 in a human intestinal cell line and a rat renal cell line [12]. We used immortalized HCECs [13] to determine if carcinine could be transported inside the cells. We first incubated HCECs with 0.2 M carcinine for 30 min, and found carcinine in the cell extract (Fig. 8.2).
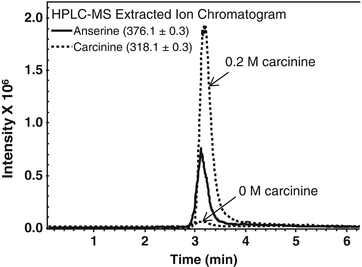
Fig. 8.2
Carcinine is found in HCECs after incubation with 0.2 M exogenous carcinine. SV-40 adenovirus immortalized HCECs were grown to 90 % confluence in 10 cm plates as previously described [13, 17, 18]. Cells were incubated with fresh basal medium containing 0 or 0.2 M carcinine for 30 min. Cells were then washed three times in PBS, harvested, and centrifuged. Cell pellet was extracted with acetonitrile, anserine was used as the internal standard, and HPLC-MS analysis of cell extracts was performed as previously described [8]. Extracted Ion Chromatogram using m/z: 318.1 ± 0.3 for carcinine and m/z: 376.1 ± 0.3 for anserine is shown. Carcinine was found in the cell extract after treatment with 0.2 M carcinine
To determine if there was an influx of carcinine in these cells, we prepared a radiolabeled 14C carcinine. 14C carcinine was synthesized according to a published procedure [14]. The identity of intermediate 14C-N-tert-butoxycarbonyl-b-alanylhistamine and final 14C-β-alanylhistamine compounds were verified by their retention times in HPLC, using unlabeled standards. In the first experiment, a time-course of carcinine accumulation was performed using a concentration of 1 mM carcinine. A fast accumulation of carcinine was observed, peaking at 5 min followed by a very slow decrease over the following 15 min (Fig. 8.3a).
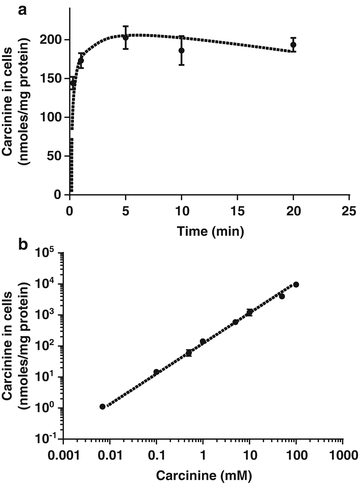
Fig. 8.3
Kinetics and dose–response of carcinine accumulation in HCECs. HCECs were grown to confluence in 24-well plates. (a) Cells were washed with basal medium once and incubated in basal medium containing 1 mM of unlabeled carcinine and a negligible concentration of 14C carcinine (0.007 mM final, 0.16 μCi/μL final) for 0, 1, 5, 10 and 20 min. Cells were washed once in PBS and harvested in 0.3 M NaOH. 14C associated with the cells was quantified in scintillation buffer. Protein concentrations were used to normalize the results. Each time point was performed in triplicate, mean and SEM were plotted. (b) Cells were washed with basal medium once and incubated in basal medium containing indicated concentrations of unlabeled carcinine and a negligible concentration of 14C carcinine (0.007 mM final, 0.16 μCi/μL final) for 20 min. Cells were washed once in PBS and harvested in 0.3 M NaOH. 14C associated with the cells was quantified in scintillation buffer. Protein concentrations were used to normalize the results. Each concentration of carcinine was performed in triplicate and error bars representing standard deviation (SD) were plotted. In most cases, they are too small to be visible on the graph
In the second experiment, we quantified the uptake of carcinine after incubation of HCECs for 20 min with concentrations of carcinine ranging from 0.01 to 100 mM. A linear dose-dependent accumulation of carcinine was found in the cells at the concentrations tested (Fig. 8.3b). This result suggests that there could be a passive, non saturable, influx of carcinine in these cells, or could indicate that carcinine simply binds the plasma membrane of the cells but does not penetrate the intracellular compartment. To determine whether or not carcinine penetrates the cells, we next investigated the biological effect of carcinine in the intracellular compartment by quantifying intracellular 4-HNE-protein adduct in HCECs.
8.3.2 Carcinine Effect in HCECs
We first incubated HCECs with 75 μM of exogenous 4-HNE for 0, 1, and 3 h, washed the cells with PBS to eliminate extracellular 4-HNE, and quantified 4-HNE-protein adduct in cell homogenates as previously described [15]. A time-dependent formation of 4-HNE-protein adduct is shown in Fig. 8.4a. We then preincubated HCECs with 0, 1, or 10 mM carcinine for 1 and 3 h, removed the media containing carcinine, washed the cells with PBS to eliminate extracellular carcinine, and then treated the cells with 75 μM of 4-HNE for 1 h. If carcinine was entering the intracellular compartment, it would inhibit the formation of 4-HNE-protein adduct in a dose-dependent manner. As shown in Fig. 8.4b, there was no significant decrease of 4-HNE-protein adduct formed in any of the cells preincubated with carcinine, suggesting that carcinine was not efficiently transported inside the cells.
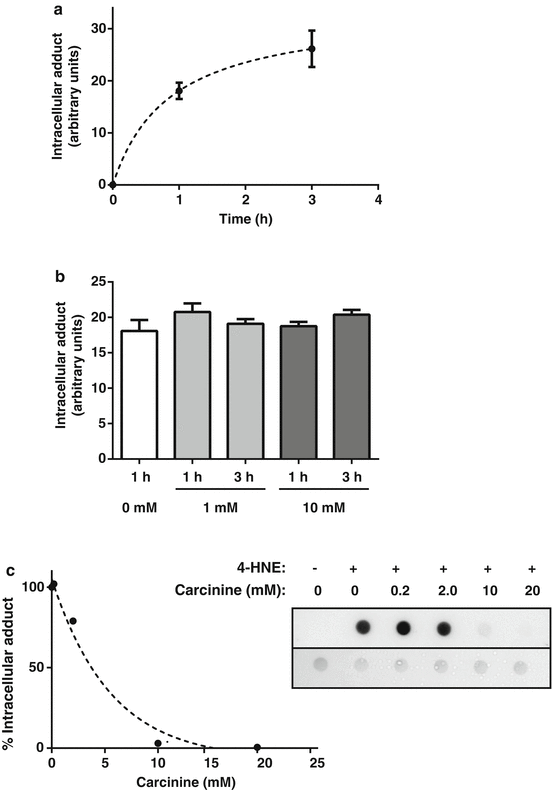
Fig. 8.4
Carcinine can quench 4-HNE in the cell medium but not in the HCEC intracellular compartment. HCECs were grown to confluence in 6-well plates. (a) Cells were washed with basal medium once and then incubated with basal medium containing 75 μM 4-HNE for 0, 1, and 3 h. (b) Cells were preincubated with basal medium containing 0, 1, or 10 mM carcinine for 1 or 3 h. At the end of the preincubation time, cells were washed three times with PBS and incubated with basal medium containing 75 μM 4-HNE for 1 h. (c) Cells were incubated with basal medium containing 75 μM 4-HNE and 0, 0.2, 2, 10 or 20 mM carcinine for 2 h. At the end of incubation times, cells were washed three times with PBS and harvested in T-PER reagent containing protease inhibitors as previously described [15]. Whole cell homogenates are prepared and equal aliquots (5 μg) of protein are analyzed by dot blot as previously described [15]. Dot blot results are quantified using ImageJ and normalized to the amount of protein in each dot quantified with Ponceau red staining of the membranes as previously described [8]. Graph shown in (a) and (b) are representative results of triplicates for each experimental condition. Means and SD are plotted. Graph shown in (c) is representative of one sample per experimental condition. The percent of intracellular adduct is plotted using 0 mM carcinine as 100 % of intracellular adduct. The dot blot showing detection of 4-HNE-protein adduct (top panel) and Ponceau staining (bottom panel) is also shown
We then treated HCECs with carcinine and 4-HNE, added simultaneously to the cell culture medium. Increasing concentrations of carcinine were used in combination with 75 μM of 4-HNE. After 2 h incubation with medium, cells were harvested and intracellular 4-HNE-protein adducts were quantified. As shown in Fig. 8.4c, carcinine completely inhibited the formation of adducts with intracellular proteins when used at concentrations of 10 mM or higher. These results show the rapid quenching effect of 4-HNE by carcinine in the cell-culture medium, inhibiting the passive influx of 4-HNE and the formation of intracellular 4-HNE-protein adduct.
Taken together, these results show that carcinine is active outside but not inside the cells, demonstrating that it does not enter the HCECs. The linear dose-dependent accumulation of carcinine found in HCEC extracts in Fig. 8.3b was probably due to an interaction of carcinine with the plasma membranes. Carcinine is a dipeptide derivative (β-alanyl-histamine), formed by a hydrophobic alanyl and a positively charged histamine. Such compound is likely to bind and be inserted in the plasma membrane but might not be able to freely cross it because of the positive charge. Our results demonstrate that no passive diffusion is taking place across cell membranes and thus suggests that the intracellular quenching effect of carcinine previously found in retinal cells [9] was probably due to a transporter-mediated uptake. The SLC15A2 mRNA has been found in the human retina [10] so it is possible that PepT2 was involved in the transport of carcinine in mouse retinal cells.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


