Chapter 33 Epigenetic Mechanisms of Retinal Disease
Brief history
The term “epigenetics” was coined by C. H. Waddington in the 1940s, fusing the word “genetics” with “epigenesis,” the latter indicating the theory by which the adult form develops from the embryo through gradual steps, as opposed to being fully preformed as a zygote.1,2 With the discovery of inheritable patterns of DNA methylation, the idea that epigenetic traits were inherited as regulatory signals in addition to genetic information quickly took hold, and the definition of epigenetics became “the study of mitotically and/or meiotically heritable changes in gene function that cannot be explained by changes in DNA sequence.” Since the 1990s, the term “epigenetics” has become commonly used to refer to heritable changes that do not involve changes in DNA sequence.3 The term “epimutation” is used to indicate a heritable change in gene expression that does not affect the actual basepair sequence of DNA. In 2003, the Human Epigenome Consortium established the Human Epigenome Project (HEP) to identify, catalog, and interpret the genomewide DNA methylation patterns of all human genes in all major tissues.4 The field of epigenetics has garnered increasing attention over recent years.
Concept
DNA methylation
DNA methylation involves the addition of a methyl group to the 5’ position of the cytosine pyrimidine ring or the number 6 nitrogen of the adenine purine ring, and typically occurs in a CpG dinucleotide context. This modification can be inherited through cell division. In mammals, 60–90% of all CpGs are methylated in the nonpromoter area.5 DNA methylation may affect the transcription of genes in two ways. First, the methylation of DNA may itself physically impede the binding of transcriptional factors to the gene, and second, and likely more important, methylated DNA may be bound by proteins known as methyl-CpG-binding domain (MBD) proteins. MBD proteins then recruit additional proteins to the locus, such as histone deacetylases (HDACs) and other chromatin remodeling proteins that can modify histones, thereby forming compact, inactive chromatin. This process has been termed silent gene expression.6
Histone methylation
Histone methylation is the modification of certain amino acids in a histone protein by the addition of one, two, or three methyl groups. Histone methylation is generally associated with transcriptional repression. However, methylation of some lysine and arginine residues of histones results in transcriptional activation.7 Histone lysine methylation has been well studied at the K4, K9, and K27 residues. These lysine residues can be monomethylated, dimethylated, or trimethylated. Generally, trimethylation of lysine 4 on histone H3 (H3K4me3) is associated with a fully activated promoter, which correlates with gene transcription, whereas dimethylation (H3K4me2) occurs at both inactive and active euchromatic genes. H3K9 is a major negative regulator of the H3K4 mark. Dimethylation at lysine 9 (H3K9me2) marks the silence of gene expression in euchromatin, important in proliferating cells, whereas H3K9me3 is enriched in regions of “gene-poor” pericentric heterochromatin. Methylation at lysine 27 on histone H3 (H3K27me) is associated with transcriptional repression in many developmental processes.8
Histone acetylation and deacetylation
In histone acetylation, the histones are acetylated on lysine residues in the N-terminal tail as an important regulatory factor of gene expression. Steady-state levels of histone acetylation are maintained by a balance between the opposing activities of histone acetyltransferases (HATs) and HDACs. Acetylation brings in a negative charge, which acts to neutralize the positive charge on the histones and decreases the interaction of the N termini of histones with the negatively charged phosphate groups of DNA. As a consequence, the condensed chromatin is transformed to a more relaxed structure, which allows transcription factors access for DNA binding and gene transcription. Aberrant histone acetylation/deacetylation and other epigenetic modulations have been implicated in many pathological conditions, including cancer and multiple sclerosis.9
Histone acetylation is catalyzed by HATs, which are broadly classified in two different classes, types A and B, based on their functional localization.9 Type A HATs are nuclear HATs that catalyze transcription-related acetylation events. Type B HATs are cytoplasmic HATs that catalyze acetylation events linked to the transport of newly synthesized histones from the cytoplasm to the nucleus for deposition on to newly replicated DNA.10 There are five main classes of HATs (GNAT, MYST, p300/CBP, transcription factor, and nuclear hormone-related), which are characterized by specific functions controlled by its specific structural folding.
The process of histone acetylation is reversed by the HDACs, which catalyze acetyl group removal.10 There are four main classes (I–IV) of HDACs. HDAC1–3, and 8 are members of class I, and are located in the nucleus and involved in epigenetic regulation. HDAC4–6, 7, 9, and 10 are members of class II, and are characterized by nucleocytoplasmic shuttling. Class III HDACs are characterized by their NAD dependence. HDAC 11 has been categorized into its own group, a class IV deacetylase. These molecules have been implicated in aging and calorie restriction as well as disease.11
Noncoding RNA
miRNAs are a class of small (on average 22 nucleotides long), noncoding RNA molecules that regulate gene expression and are implicated in many cellular processes, including development, tissue morphogenesis, apoptosis, and tumor growth.12–14 Genes encoding primary miRNAs are scattered throughout the genomes of eukaryotes and are transcribed to generate RNA species that are cleaved to 60–70-nucleotide stem–loop miRNA precursors (pre-miRNAs) by a microprocessor complex containing the nuclear RNaseIII Drosha and DiGeorge syndrome critical region protein. After export from the nucleus, the pre-miRNAs are cleaved by a cytoplasmic RNase III (Dicer). After strand selection and separation, mature 22-basepair miRNAs are incorporated into an RNA-induced silencing complex to act as target recognition sequences. miRNAs may contain target sequences and can undergo cleavage or translational repression.15
Epigenetic factors in the retina
DNA methylation in mammalian retina
DNA methyltransferases (DNMT) are expressed in mammalian retina. Three active DNMTs have been identified in mammals: DNMT1, DNMT3a, and DNMT3b. DNMT3a methylates CpG sites at a rate much slower than that of DNMT1 but greater than that of DNMT3b. DMNT1 is expressed in human retinal pigment epithelium (RPE) cells. DNMT3a is weakly expressed in some inner nuclear layer cells in the adult mouse retina.16
DNA methylation regulates the expression of genes in mammalian retina. Interphotoreceptor retinoid-binding protein (IRBP) is the major soluble component of the interphotoreceptor matrix. This important extracellular matrix (ECM) surrounds the outer and inner segments of the photoreceptors and separates the neural retina from the RPE. IRBP is a large glycolipoprotein, which is secreted into this matrix by both rods and cones. The IRBP gene is hypomethylated in retina, and DNA methylation regulates IRBP gene expression.17 Exogenous methylation of specific CCGG sites in the murine IRBP 5’ flanking region suppressed promoter activity in transient transfection assays of cultured retinal cells. Methylation of the IRBP promoter suppresses transcription in nonphotoreceptor cells by precluding specific DNA:protein-binding events, whereas the lack of methylation in photoreceptors allows for transcription.17 Another example is EphA5, a member of the ephrin receptor subfamily of the protein-tyrosine kinase family. It regulates multiple aspects of development and disease, including vascular development. In the retina of the adult mouse, EphA5 mediates 1.2 ± 0.3% of CpG methylation.18 This represents a relatively low level of promoter methylation. Although methylation of all CpG sites resulted in the silencing of EphA5 promoter activity, lower levels of methylation resulted in differential activation or repression of EphA5 promoter activity, depending on the sites methylated.18 We found that methyl CpG-binding protein 2 (MeCP2) and DNMT1 are expressed in human RPE cells.
Histone methylation and acetylation in mammalian retina
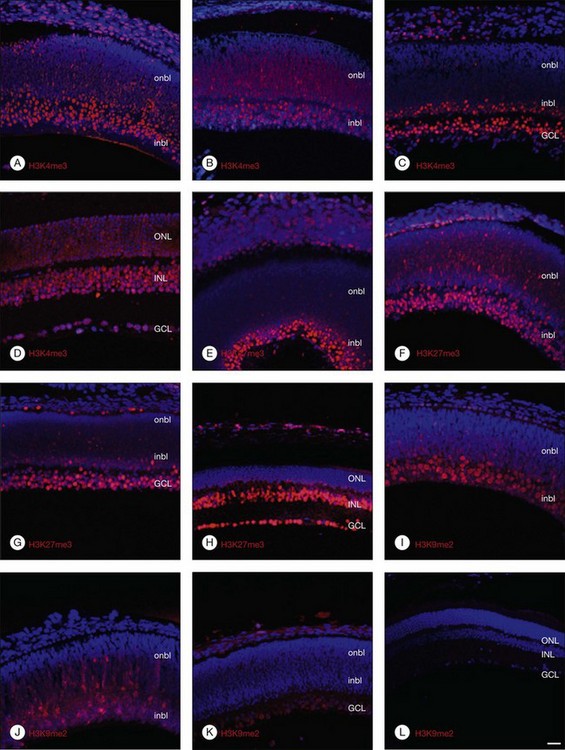
Histone lysine methylation is catalyzed by histone methyltransferases (HMTases). The HMTases constitute a family of enzymes that catalyze the methylation of histones on specific residues. The HMTases have been observed in mammalian retina. In the adult murine retina, H3K4me3, a mark associated with active transcription, is observed in all layers of the neural retina, including rhodopsin-positive photoreceptors, Müller glia, and retinal ganglion cells (RGCs). H3K27me3, a mark associated with transcriptional repression, is enriched in the inner nuclear layers and in a subset of outer nuclear layer in the adult murine retina. However, H3K9me2, another repressive mark, is not observed in the inner layers of the adult retina.8 The Ezh2 and G9a HMTases are the two best-characterized HMTases, which catalyze H3K27me3 and H3K9me2 modifications, respectively. Both can be found in the fetal murine retina. The results suggest that epigenetic regulation of gene transcription by Ezh2- and G9a-mediated histone lysine methylation plays a crucial role in retinal neuron differentiation and survival.8
HDAC is expressed in mammalian retina.19 Acetylated histone 3 and 4 are found in fetal and adult retina; however, acetylated histone 3 and 4 is reduced in dry age-related macular degeneration (AMD) specimens. In contrast to the decrease of acetylated histones, HDAC1 is highly upregulated in human AMD retinal sections. In addition, exposure of adult mice retina to trichostatin A (TSA) induces upregulation of apoptotic genes. Activation of HDAC may decrease the retina’s resistance to ischemic injury.20
miRNA in mammalian retina
The expression of miRNAs in the mammalian retina has been analyzed in several studies. The most highly expressed adult mouse retinal miRNAs were miR-181a, 182, 183, 204, 125b, 26a, and 124a. miR-181 is expressed in the ganglion cell, inner plexiform and inner nuclear layers; miR182 is expressed throughout all layers of the retina; and miR-183 is expressed only in the outer nuclear layer.21 Another study found strong expression of miR-204 in the RPE and the ciliary body, and miR-224 was detected in the RPE. In contrast, miR-124a was expressed in all layers of the neural retina except the RPE. Most recently, miRNA-132 was found to be expressed in human RPE cells. miR-204 and 211 were also found to be highly expressed in the RPE. Transforming growth factor-β (TGF-β) receptor-2 and SNAIL2 are direct targets of miR-204. Notably, anti-miR-204/211 decreased transepithelial resistance and reduced cell membrane voltage and conductance, indicating a critical role of miR-204/211 in maintaining epithelial barrier function and cell physiology.22
Epigenetic mechanisms in retinal development
Although epigenetic mechanisms have been shown to regulate neural stem cell renewal and differentiation, it was not until recent years that their involvement in retinal development has been realized. The retina is composed of specialized glia and neuronal cells which are generated from multipotent retinal progenitor cells in a highly conserved temporal sequence with overlapping phases during development.23,24 RGCs, horizontal cells, and cone photoreceptors are born in early phases, while rod photoreceptors, bipolar cells, and Müller glial cells are born in late phases. The cell fate decision of the progenitor cells depends on both intrinsic and environmental cues, which are regulated by specific networks of transcription factors.23,24 Through covalent modifications of DNA and histones, chromatin remodeling regulates the interactions of these transcription factors and their effector genes, and is an important epigenetic mechanism in retinal development.
DNA and histone methylation in retinal development
DNA and histone methylation are mediated by DNMT and HMTases. In zebrafish retina, antisense-based morpholino knockdown of DNMT3 and H3K9 HMTases Suv39h1 and G9a lead to defects in retinal cell differentiation, supporting a role of both DNA and histone methylation in zebrafish retinal development.25 In mice, immunohistochemical analysis shows changing patterns of histone methylation markers in the developing retina. Specifically, the transcriptionally activating H3K4me3 and repressive H3K27me3 histone marks are found in differentiated neurons from embryo to adulthood, corresponding to the expression of the HMTase Ezh2 that catalyzes the H3K27me3 mark. In contrast, the repressive H3K9me2 mark and the corresponding HMTase G9a were found primarily in early differentiating RGCs, but decreased after birth (Fig. 33.1).8 These changing patterns of histone methylation may at least in part account for the temporal sequence of retinal progenitor cell differentiation during development. Evidence also implicates DNA methylation in regulating the expression of genes involved in the topographic patterning of RGC axons, such as EphA5 receptor.18 These studies suggest that DNA and histone modifications may regulate both temporal and spatial expression of these genes in different populations of retinal progenitor cells to orchestrate the precise timing of cell proliferation and differentiation during development.
Histone acetylation in retinal development
During photoreceptor differentiation, HAT-containing coactivators, such as general control nondepressible 5 (Gcn5) and CREB-binding protein (Cbp), interact with cone–rod homeobox (Crx) transcription factor to promote the transcription of opsin genes.26 Crx-deficient mutant mice have decreased histone acetylation at the promoter/enhancer region of the opsin gene, resulting in photoreceptor dysfunction.27 Such results indicate that HATs are required for differentiation of retinal photoreceptors during development.
In addition to HATs, HDACs have been studied more extensively and are also implicated in retinal development. HDAC1 is recruited by the retinoblastoma protein (Rb) and related family members to promoters with E2F-binding sites, resulting in transcriptional repression of cell cycle genes and suppression of cell growth. In zebrafish, hdac1-deficient mutants exhibit retinal cell proliferation and differentiation defects, resulting in severe reduction in the inner and outer plexiform layers and absence of RGC, rod and cone photoreceptors, and Müller glia.27 These defects result from failure in downregulation of cyclin D and E transcription and the canonical Wnt and Notch signaling pathways, which are necessary for retinal progenitor cells to exit the cell cycle. Thus, histone deacetylation in zebrafish may be a regulatory mechanism to switch off stem cell proliferation and initiate a program for retinal differentiation. In mouse retinal explants, pharmacological inhibition of HDAC with TSA also results in defects in rod differentiation, but unlike in zebrafish, causes a reduction in retinal progenitor cell proliferation. In vivo knockdown HDAC4 by RNA interference leads to apoptosis of rod photoreceptors and bipolar cells, while overexpression of HDAC4 reduced naturally occurring cell death of bipolar cells, supporting a role of HDAC4 in promoting the survival of developing retinal neurons as well.28 Further work will help clarify what specific downstream effectors are regulated by HATs and HDACs during the various stages of retinal development.
Chromatin remodeling complexes in retinal development
While covalent DNA and histone modifications decondense chromatin and promoter regions, ATP-dependent displacement from promoters and enhancer regions requires the action of chromatin-remodeling complexes. These include the SWItch/Sucrose NonFermentable (SWI/SNF) and the Imitation SWItch (ISWI) families. Zebrafish mutants lacking the brahma-related gene 1 (BRG1), the catalytic ATPase subunit of SWI/SNF chromatin remodeling complex, show defects in retinal differentiation, suggesting that BRG1 is involved in triggering retinal cell differentiation.29 In mice, however, loss of BRG1 in neural stem cells results in precocious neuronal differentiation, suggesting that it represses differentiation and maintains neural stem and progenitor cells. These differences may be related to changes in the composition of accessory units known as BRG1-associated factors (BAFs) in the SWI/SNF complexes in different cell states. Nevertheless, chromatin-remodeling complexes are likely to be critically involved in regulating retinal cell differentiation during development.
microRNAs in retinal development
Recently, miRNAs have also been implicated in retinal development. Early studies using a conditional knockout mouse lacking the RNA endonuclease Dicer in the retina showed no defects in the retina before the second postnatal week, suggesting that miRNAs are not required for mouse retinal development.30 However, subsequent research using a different strain of Dicer conditional knockout mouse showed increased production of early generated cell types such as RGC and horizontal cells, and failure to generate late-born cell types such as rod photoreceptors and Müller glia.31 This is also supported by data from Xenopus morpholinos, where Dicer inactivation resulted in defects in cell cycle, lamination, and timing of retinal differentiation. These observations suggest that Dicer and miRNAs may provide a common regulatory mechanism to signal changes in retinal progenitor cell competence.
Microarray analysis of the miRNA transcriptome in mouse retina reveals at least 78 miRNAs, 21 of which are potentially retina-specific.32 The miRNA transcriptome of the mouse retina is similar to that of humans, and shows dozens of miRNAs that are differentially expressed during different stages of development. In Xenopus, miRNAs have been found to inhibit the translation of homeodomain transcription factors (Xotx2 and Xvsx1) involved in bipolar cell differentiation by binding the 3’ untranslated region of mRNAs. Inactivation of these miRNAs in vivo releases the inhibition and supports the generation of additional bipolar cells.33 Another miRNA, miR-24a, negatively regulates proapoptotic factors caspase-9 and apoptotic protease-activating factor 1 (apaf1) in Xenopus retina. Inhibition of miR-24a leads to increased apoptosis during retina development and reduction in eye size.34 These studies suggest that specific miRNAs can play distinct roles in signaling differentiation or promoting survival of retinal neurons. In mice, the miRNAs most enriched in the retina include miR-96, miR-182, and miR-183, which are transcribed as a single precursor pri-miRNA.32 Target prediction and in vitro functional studies showed that the microphthalmia-associated transcription factor (MITF), a transcription factor necessary for the development and function of RPE, is directly inhibited by miR-96 and miR-182, supporting a role of miRNAs in RPE maintenance also.32 MiR-124a, which is expressed in all neuronal subtypes in the adult retina, has also been shown to repress retinol dehydrogenase 10 (Rdh10), which is selectively expressed in Müller glia and RPE.35 Hence, some miRNAs may impact retinal development not by affecting neuronal proliferation and differentiation, but via effects on supporting glial and RPE cells. MicroRNA research is still in its infancy, and more work is necessary to determine the landscape of noncoding RNAs in different retinal cell populations.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


