CHAPTER 128 Anatomy of the Auditory System
Cochlear Anatomy
Osteology
The inner ear is contained in the petrous apex of the temporal bone, and is encased in a bony structure called the osseous or bony labyrinth. The bone forming the cochlea and vestibular labyrinth is the hardest bone in the human body and is akin to ivory in its density. The labyrinth consists of three continuous sections: the vestibule, the cochlea, and the semicircular canals. The initial point of communication between the middle and inner ears occurs at the oval window of the vestibule, where the stapes footplate abuts the oval window membrane. At the basal end of the cochlea is the round window membrane, which communicates with the middle ear space. The cochlea is a snail-shaped structure that has a wide diameter at the base, which narrows for 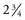 turns until it reaches its apex. Figure 128-1 shows a cross section of the cochlea, illustrating the cuts through each turn.
turns until it reaches its apex. Figure 128-1 shows a cross section of the cochlea, illustrating the cuts through each turn.
Membranous Labyrinth and Inner Ear Fluids
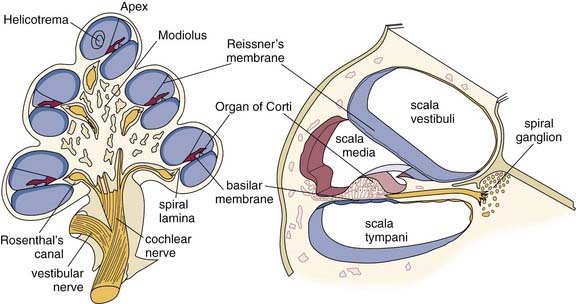
The membranous labyrinth of the cochlea follows the shape of the osseous cochlea and forms a third cochlear chamber—the scala media. The sensory organ of hearing resides within the membranous labyrinth. Figure 128-2 shows a cross section of the membranous labyrinth and its structures. The membranous labyrinth is bordered superiorly by Reissner’s membrane, inferiorly by the basilar membrane, and laterally by a portion of the outer cochlear wall to which it is anchored by the spiral ligament. Within the membranous labyrinth, along the lateral wall, is the stria vascularis, highly vascular tissue that is responsible for the metabolic environment of the scala media. The intricate structures that comprise the organ of Corti are situated on the basilar membrane.
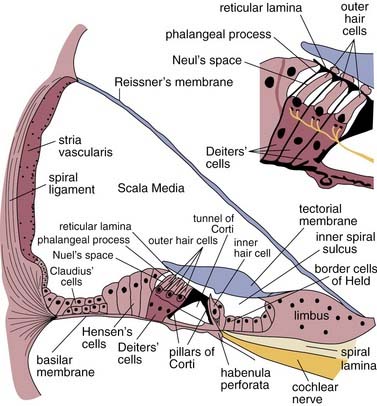
The organ of Corti runs longitudinally along the length of the basilar membrane and consists of many types of epithelial cells and structures. Medially, seated atop the osseous spiral lamina is the spiral limbus, a thickened band of periosteum that serves as the point of medial attachment for Reissner’s membrane and gives rise to the tectorial membrane. The tectorial membrane lies over the inner and outer hair cells. It is a compliant gelatinous structure composed primarily of collagen II fibers, and serves as a mass load that moves similar to a rubber band.1–3 Lateral to the spiral limbus is the inner spiral sulcus, which is lined with the border cells of Held. One row of inner hair cells is present, and the cell bodies are surrounded by supporting cells called phalangeal cells.
The two fluid systems within the cochlea create an environment crucial to the mechanical displacement of the basilar membrane traveling wave and to the cellular depolarization and subsequent synaptic activity. Between the osseous and membranous labyrinths is perilymphatic fluid, or perilymph, which has a high concentration of sodium and low concentration of potassium, similar to what is found in cerebrospinal fluid and blood serum. The presence of perilymph in the intercellular spaces within the organ of Corti has been identified in humans and guinea pigs.4,5
Within the membranous labyrinth is endolymphatic fluid, or endolymph, with a high concentration of potassium and low concentration of sodium, such as is typically found intracellularly. The ionic concentrations of endolymph are maintained by the cells within the stria vascularis. The endolymphatic sac communicates with the membranous labyrinth via the endolymphatic duct and vestibular aqueduct. Disorders of the endolymphatic system resulting from disease, congenital deformities, or other insults can lead to potentially severe auditory and vestibular symptoms. An enlarged vestibular aqueduct, defined as a diameter larger than 1.5 mm at the vestibular aqueduct midpoint or greater than 2 mm at the operculum, may lead to sudden and progressive sensorineural hearing loss in children, particularly after even minor head trauma.6
Hair Cells
The inner and outer hair cells function as receptor cells that transduce mechanical movement into an electrochemical signal to stimulate the auditory nerve. Figure 128-3 presents schematic examples of inner and outer hair cells. The apical portion of all hair cells includes a thickened region called the cuticular plate, which in conjunction with the supporting cells forms the reticular lamina. Rooted in the cuticular plate of each hair cell and projecting through the reticular lamina are bundles of actin filaments called stereocilia, stiff hairlike structures that deflect with mechanical disturbances. Adjacent to this is a noncuticular region that contains a rudimentary kinocilium.
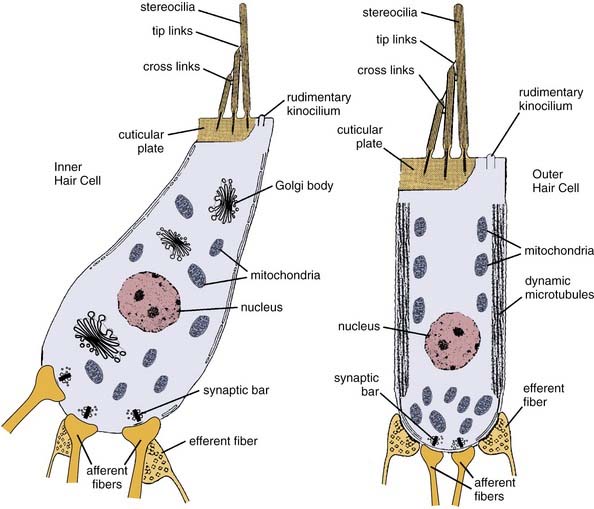
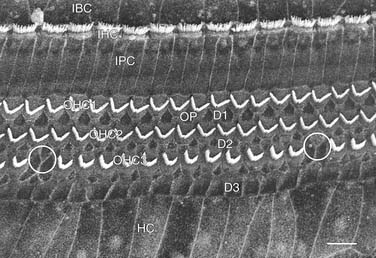
There are approximately 3500 inner hair cells arranged in one row on the modiolar side of the tunnel of Corti and 12,000 outer hair cells in three rows on the strial side. Figure 128-4 is a scanning electron micrograph of the reticular lamina of the organ of Corti showing the stereocilia arrangement for the three rows of outer hair cells and single row of inner hair cells. The stereocilia on outer hair cells form a “V” or “W” shape with the bottom facing away from the modiolus. The inner hair cell stereocilia form a shallow “U” that opens toward the modiolus. On each hair cell, there are rows of stereocilia, two or more on inner hair cells, and three or more on outer hair cells. For both types of hair cells, the stereocilia lengths are graduated from longest on the strial side to shortest on the modiolar side. The longest stereocilia on the outer hair cells contact the tectorial membrane, which results in deflection of the stereocilia with basilar membrane movement. The stereocilia are connected to each other by filamentous links laterally by cross-links and from the tips of shorter stereocilia to the sides of the taller ones by tip-links. These ensure that the connected stereocilia move as a unit when the longer stereocilia are deflected.
The structural aspects of the inner and outer hair cell bodies themselves differ significantly and are reflective of their functional differences. The inner hair cells are flask-shaped, wide at the bottom and narrower at the top, and contain high concentrations of organelles that are involved in metabolic activity, particularly Golgi bodies and mitochondria. Although highly metabolic, inner hair cells are considered to be passive transducers in the auditory system. Outer hair cells are cylindrical in shape and contain microfilaments and microtubules along the length of the cell that give rise to motile activity.7 The motile properties have been shown empirically to result in highly tuned, frequency-specific contractile activity even when stimulated in isolation from the basilar membrane.8 All hair cells have synaptic bars at afferent synapses, which serve as sites for presynaptic vesicle docking and release for subsequent stimulation of auditory nerve fibers.
Innervation
Afferent innervation of the cochlear hair cells consists of approximately 30,000 auditory nerve fibers in humans, and is responsible for providing ascending information from the cochlea to the central auditory system. The cell bodies of afferent fibers make up the spiral ganglion that resides in Rosenthal’s canal within the modiolus. Nerve fibers reach the hair cells by traveling through the modiolus, and into the osseous spiral lamina, where they pass through holes in the lamina referred to as the habenulae perforatae. The nerve fibers are classified as type I and type II. Type I fibers are bipolar, of large diameter, and myelinated, and they constitute nearly 95% of the total number of fibers. Each type I fiber has a direct and independent synapse on the body of a single inner hair cell, and each inner hair cell is innervated by approximately 20 such fibers.9,10 Type II fibers constitute the remaining 5%, and are smaller and may be myelinated or unmyelinated. Type II fibers synapse directly on the outer hair cells, with a single fiber diverging to form branches that synapse with multiple other outer hair cells.
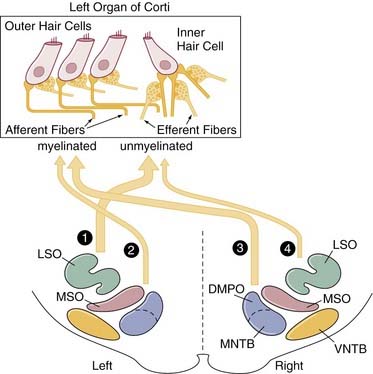
The efferent auditory pathway originates from the olivocochlear bundle and provides central inhibitory modulation of hair cell activity via descending information. The olivocochlear bundle has about 1600 fibers, and these constitute the uncrossed olivocochlear bundle and crossed olivocochlear bundle.11–15 These pathways originate from the medial superior olive (MSO) and lateral superior olive (LSO) regions on both sides. For the uncrossed olivocochlear bundle, the LSO projects many small-diameter, unmyelinated efferent fibers toward the ipsilateral inner hair cells, where they synapse with the afferent fibers. The MSO projects fewer myelinated fibers ipsilaterally that synapse directly on the outer hair cells. For the crossed olivocochlear bundle, the MSO projects large-diameter myelinated fibers to the contralateral outer hair cells, and the LSO projects a few unmyelinated fibers to the contralateral inner hair cell afferents. The fibers from the crossed olivocochlear bundle cross midline at the level of the fourth ventricle. To illustrate the pathways, Figure 128-5 shows a schematic example of the uncrossed olivocochlear bundle and crossed olivocochlear bundle innervation for the left organ of Corti.