(1)
Department of Ophthalmology, St. Thomas’ Hospital, London, UK
1.1.1 The Vitreous
1.1.2 The Retina
1.2.1 Using the Database
1.2.2 Examination of the Eye
1.2.3 Subjective Tests
1.3 Summary
Abstract
During early development, the invaginated optic vesicle (optic cup) contains the primary vitreous, a vascularised tissue supplying the lens and retina (both of which have an ectodermal origin). During the third month of gestation, the primary vitreous gradually loses its vascularity and is replaced by the secondary vitreous derived mainly from the anterior retina and ciliary body. The principal remnants of the primary vitreous are Cloquet’s canal and some epipapillary gliosis. A mild exaggeration of the latter is seen in Bergmeister’s papilla (fibrous tuft) on the optic nerve head, whilst a Mittendorf’s dot is a primary vitreous remnant on the posterior capsule of the lens. The hyaloid artery may occasionally persist as a vascular channel growing into the central gel from the optic disc or as a glial plaque on the posterior lens capsule (see Chap. 13).
1.1 Surgical Anatomy of the Retina and Vitreous
1.1.1 The Vitreous
Embryology
During early development, the invaginated optic vesicle (optic cup) contains the primary vitreous, a vascularised tissue supplying the lens and retina (both of which have an ectodermal origin). During the third month of gestation, the primary vitreous gradually loses its vascularity and is replaced by the secondary vitreous derived mainly from the anterior retina and ciliary body. The principal remnants of the primary vitreous are Cloquet’s canal and some epipapillary gliosis. A mild exaggeration of the latter is seen in Bergmeister’s papilla (fibrous tuft) on the optic nerve head, whilst a Mittendorf’s dot is a primary vitreous remnant on the posterior capsule of the lens. The hyaloid artery may occasionally persist as a vascular channel growing into the central gel from the optic disc or as a glial plaque on the posterior lens capsule (see Chap. 13).
The Anatomy
The vitreous cavity is the space within the eye bounded anteriorly by the lens and its zonular fibres, and more posteriorly by the ciliary body, retina and optic disc. Its volume is usually about 4 ml, though this may increase to as much as 10 ml in highly myopic eyes. Normally, the space is entirely occupied by vitreous gel, a virtually acellular viscous fluid with 99 % water content. Its low molecular and cellular content is essential for the maintenance of transparency. The major molecular constituents of the vitreous gel are hyaluronic acid and type 2 collagen fibrils. The cortical part of the vitreous gel has a higher content of hyaluronic acid and collagen compared with the less dense central gel. In addition, the gel exhibits ‘condensations’ both within its substance and along its boundaries. The boundary condensations are termed the anterior and posterior hyaloid membranes. A central tubular condensation called Cloquet’s canal is a remnant of the primary vitreous, stretching sinuously between the lens anteriorly and the optic disc posteriorly. The gel is unimportant in maintaining the shape or structure of the eye. Indeed, apart from its role in oculogenesis, the vitreous has no well-substantiated function. An eye devoid of gel is not adversely affected apart from a poorly understood increased risk of nuclear sclerotic cataract. The pO2 of the vitreous is relatively low, and it has been suggested that the vitreous may act to reduce oxidative stress on the lens fibres thereby reducing cataractogenesis (Stefansson et al. 1982). The vitreous gel is however primarily implicated in the pathogenesis of a variety of sight-threatening conditions.
Anatomical Attachments of the Vitreous to the Surrounding Structures
The posterior hyaloid membrane adheres to the internal limiting membrane of the retina by the insertion of vitreous gel fibrils. The internal limiting membrane has type 4 collagen and is the basement membrane of the Muller cells. The potential space between the internal limiting membrane and the posterior hyaloid membrane is the plane of cleavage of the gel from the retina in posterior vitreous detachment.
The vitreous possesses various sites of increased adhesion to the surrounding structures. These attachments form the basis of much vitreoretinal pathology.
The vitreous base is an annular zone of adhesion some 3–4 mm wide which straddles the ora serrata. Its anterior border is the site of insertion of the anterior hyaloid membrane. The posterior border of the vitreous base is surgically important because this is the anterior limit of potential separation between the gel and the retina and a common site for retinal tear formation. Adhesion of the vitreous base to the retina and the pars plana is difficult to break even with severe trauma.
Weigert’s ligament is a circular zone of adhesion, 8–9 mm in diameter, between the gel and the posterior lens capsule. It is the junction between the anterior hyaloid membrane and the expanded anterior portion of Cloquet’s canal.
The posterior hyaloid membrane and the slightly expanded posterior limit of Cloquet’s canal meet around the margin of the optic disc and produce another ring of adhesion. During posterior vitreous detachment, gliotic tissue is avulsed from the edge of the nerve head to produce the Weiss ring which can be used as an indicator of posterior vitreous detachment (see Fig. 1.1). A circle of relatively increased adhesion to the retina may be present in the parafoveal area and implicated in macular hole formation.
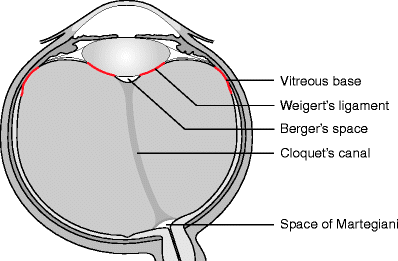
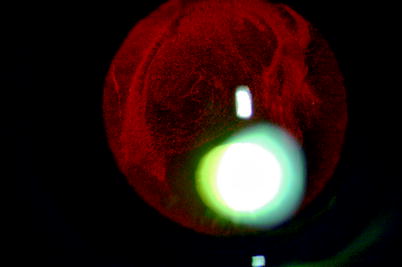
Fig. 1.1
The macroscopic anatomical landmarks are shown
Fig. 1.2
Using retroillumination, Weigert’s ligament on the back of the lens can be seen in this patient with mild vitreous haemorrhage
Exaggerated vitreoretinal adhesions are also present in lattice degeneration which comprises oval or elongated areas of thinning and vascular sclerosis in the peripheral retina with overlying degenerative vitreous gel. The lesions are generally orientated circumferentially but may be radially directed along the post-equatorial course of retinal veins. Lattice degeneration is found in approximately 7 % of normal eyes and is frequently associated with tearing of the retina. The surgeon can experience the adhesion of the vitreous to lattice during induced vitreous separation in macular hole surgery (see Chap. 8). Trying to pull the vitreous off lattice will result in tearing of the retina. Some eyes also demonstrate abnormally strong vitreoretinal adhesions along the course of the retinal veins (paravascular adhesions) which may result in retinal tear formation.
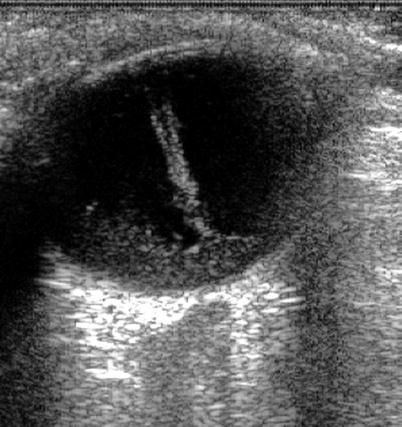
Fig. 1.3
An ultrasound of vitreous haemorrhage (seen especially in the retrohyaloid space) shows Cloquet’s canal delineated by the haemorrhage
1.1.2 The Retina
Embryology
The optic cup develops from the optic vesicle at 6–7 weeks of gestation and consists of two layers of ectoderm, the outer becoming the retinal pigment epithelium and the inner the neurosensory retina. The space between the two layers is the same as the ‘subretinal’ space in retinal detachment. The retina develops as two neuroblastic layers, inner and outer, which differentiate into the various cells of the retina. The receptor cells are the last to appear at approximately 13 weeks. At 5.5 months, the adult arrangement can be seen, but the retina is not completely developed until 3–4 months after birth when the macula is formed. The retinal pigment epithelium becomes pigmented from 6 weeks to 3 months of gestation.
Anatomy
The retina is divided into regions with the macula consisting of the area between the temporal vascular arcades, serving approximately 20° of visual field. The fovea is the central darkened area with a pit called the foveola. The cones are densest at the fovea, at 15,000/mm2, with 4,000–5,000/mm2 in the macula. There are 6 million cones and 120 million rods in total.
Cones provide high-resolution colour vision in photopic conditions. They react quickly and recover rapidly to different light stimuli. Three types of cone photoreceptor exist in the human eye with different opsin proteins bound to a common chromophore (11-cis-retinal). The three types provide sensitivities which peak at different light wavelengths: short S cones at 420 nm (blue), middle M cones at 530 nm (green) and long L cones at 560 nm (red).
The retina is organised into four layers of cells and two layers of neuronal connection. The retina has a structural cell called the Muller cell which extends through all the layers. These cells are specialised glial cells which hold a sink of ions during depolarisation of receptors and are essential for the physiology of the eye. They may also have functions in:
Cone neuroprotection
Control of vascular permeability and haemostasis
Pigment recycling
There are astrocytes and microglial cells in addition in the retina.
The retinal layers, from outer to inner retina, are as follows:
Retinal Pigment Epithelium (RPE)
A single layer of pigmented cuboidal epithelial cells which look after the function of the receptors by:
Absorbing stray light (using melanin pigment)
Transporting metabolites between the receptors and the choroid
Providing a blood–retinal barrier
Regenerating the visual pigments
Phagocytosing the receptor outer segments leading to lipofuscin production
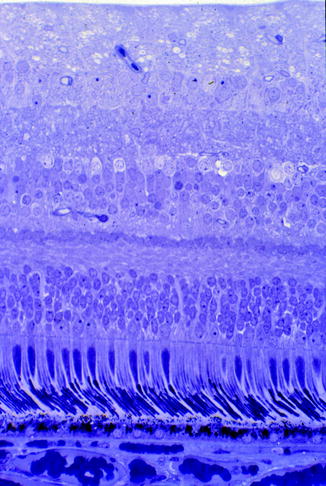
Fig. 1.4
The foveal anatomy showing increased numbers of cones and absence of the nerve fibre layer. (Courtesy of John Marshall)
Fig. 1.5
The normal stratified structure of the retina (Courtesy of John Marshall)
Photoreceptor Layer
The photoreceptor transduces light into neuronal signals. The action of light closes gated cation channels leading to hyperpolarisation of the cell. Two types of photoreceptor exist, the rods predominantly in the periphery and absent from the fovea and the cones concentrated at the macula. They are made up of:
Outer Segments
Light is absorbed by the visual pigments which are contained in stacked discs. The discs are separate in the rods (1,000 in number) but are interconnected in the cones. These join to the inner segment by the cilium.
Inner Segments
These consist of an inner myoid which contains the Golgi apparatus and ribosomes for making cell structures and an outer ellipsoid which contains mitochondria for energy production. These connect to the nucleus by the outer connecting fibre. The inner connecting fibre connects to the synaptic region. The latter has synapses arranged as triads with connections to one bipolar cell and two horizontal cells. In cones, there may be up to 20 triads whereas the rods have only one.
Outer Limiting Layer
This consists of junctional complexes from the Muller cells and photoreceptors and is located at the inner connecting fibres.
Outer Plexiform Layer
The cell processes of the horizontal cells and bipolar cells synapse with the receptors.
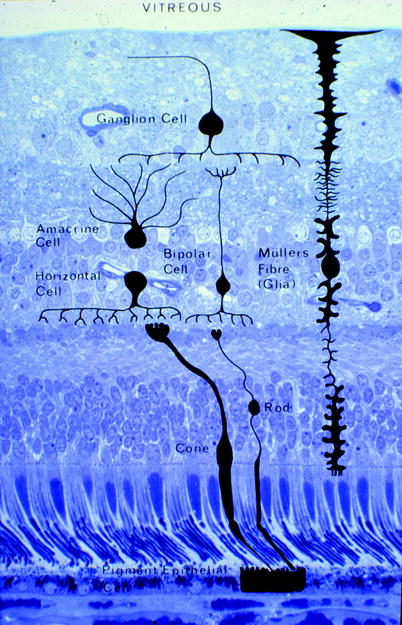
Fig. 1.6
The cell types are shown, and their positions in the retina are indicated (Courtesy of John Marshall)
Intermediary Neurones
Inner Nuclear Layer
This contains the cell bodies of the bipolar cells, the Muller cells, amacrine cells and horizontal cells.
Inner Plexiform Layer
The bipolar cells axons pass through, synapsing with the amacrine cells which help process the neuronal signals to the ganglion cells.
Ganglion Cells
Ganglion Cell Layer
The cell bodies of the ganglion cells are found here. These cells have gathered preprocessed information from the other retinal cells. The cells receive different visual information such as a sustained response to light, transient response or response to movement. At the macula, there is 1 ganglion cell to 1 receptor, but on average, in the whole retina, there is 1 for 130 receptors.
Nerve Fibre Layer
The nerve fibres of the ganglion cells on the inner surface of the retina pass tangentially towards the optic nerve.
Inner Limiting Membrane
Note: The internal limiting membrane (ILM) is a tough membrane laid down by the Muller cells with connections to the hyaloid membrane of the vitreous.
Retinal Blood Vessels
The central retinal artery supplies the neural retina with the exception of the photoreceptors which are supplied by the choriocapillaris. This is an end artery system with a single draining vessel, the central retinal vein. Both vessels have four main branches which divide at the optic disc to supply nasal and temporal quadrants. At the posterior pole, there is a capillary network at the level of the nerve fibre layer and the outer plexiform layer. In the periphery, there is one capillary network at the inner nuclear layer. The capillary endothelium forms the inner retinal blood–retinal barrier by having tight intercellular junctions.
Other Fundal Structures
Bruch’s Membrane
A pentalaminar structure partly representing the basement membranes of the RPE and the choriocapillaris. It is of ectodermal and mesodermal origin. Accumulation of damage in Bruch’s is seen in age-related macular degeneration.
Choroid
This is a vascular layer (large vessels are outer and the capillaries are inner) with a high relative blood flow and low oxygen utilisation (3 %). It supplies the RPE and photoreceptors. The highly anastomotic and fenestrated capillaries are arranged into lobules and are supplied by the posterior ciliary arteries and drained by the vortex veins.
1.1.3 The Physiology of the Vitreous
The physiology of the vitreous is not well understood. It is thought that molecules can move in the gel because of diffusion and convection and by the effects of saccades on the fluid component of the gel. Diffusion is most important for animals with smaller eyes, whereas convection is more important for larger eyes such as human eyes. Molecules with an anionic charge move more easily in the gel. Small soluble molecules like fluorescein move at a similar rate to aqueous; larger molecules like albumin may move 30–50 % less rapidly than aqueous (Xu et al. 2000). Convection is estimated to account for 30 % of movement of molecules in the vitreous because there is a pressure differential from the anterior ingress of aqueous to the posterior egress of fluid through the retina by the RPE pump. The saccadic movement of the eyes induces convection currents in the anterior vitreous which circulate around the vitreous base because of the effect of the indentation of the lens into the vitreous cavity (Repetto et al. 2010).
The measured viscosity of the vitreous depends on the technique used with estimates saying from 5–2,000 mPas (aqueous = 1 mPas). It is non-Newtonian, that is, a non-linear relationship and bimodal probably because there is a micro- and macro-viscosity component. The vitreous is relatively hypoxic (pO2 of 30–40 mmHg) in comparison to air (150 mmHg) and arterial blood (100 mmHg) (Stefansson 2006). The relatively small proportion of ocular blood supply to the retina (2–3 %) has a profound effect on the vitreal PO2 (Williamson and Harris 1994; Shui et al. 2009). There are oxygen gradients in the vitreous with higher PO2 in the anterior vitreous than the posterior. Ascorbate concentrations are high in the vitreous; the ascorbate may react with oxygen to reduce the PO2 (Shui et al. 2009).
The reason for low PO2 in the vitreous is unknown, but it may be to protect the lens proteins from oxidation (Holekamp et al. 2005). Removal of the vitreous by vitrectomy causes nuclear sclerotic cataract except in ischemic eyes such as diabetic retinopathy and retinal vein occlusion (Holekamp et al. 2006). Inserting tamponade agents such as silicone oil into the vitreous cavity increases the concentration of ions, for example, K+ and Ca+ in the remaining aqueous layer in the cavity (Winter et al. 2000).
1.1.4 Anatomy and Physiology and the Vitreoretinal Surgeon
There are certain features of the anatomy and physiology that the surgeon should remember whilst operating.
At the ora serrata, the non-pigmented epithelium is continuous with the neurosensory retina, and, therefore, retinal detachments can extend anteriorly through the ora on rare occasions. Ultimately, the ciliary body may be detached causing reduction in intraocular pressure (IOP) and even hypotony and choroidal effusion.
The posterior attachment of the vitreous base to the retina moves more posteriorly in the elderly (Bishop et al. 2004).
The nerve fibre layer orientation is especially important whilst working on the surface of the retina where damage to the nerve fibres might occur, for example, in macular hole surgery (Chap. 8).
The fovea is the thinnest part of the retina and is therefore prone to dehiscence and hole formation during retinal elevation, for example, in macular surgery.
The force required to cause a retinal detachment has been put at approximately 200 dynes/cm2 (approximately 0.27 mmHg, i.e. not very much) in primates. A number of mechanisms have been implicated in keeping the retina attached.
The retinal pigment epithelium applies forces to the retina: through ionic flow as calcium and magnesium move across the RPE, hydrostatic forces from the intraocular pressure and flow of fluid out of the eye. The RPE pump works against the relative resistance to fluid flow of the retina and has been estimated at 0.3 ml/h/mm2.
Increased osmotic pressure exerted by the increased protein content in the choroidal circulation also encourages fluid flow across the retina.
In addition, there are intercellular components aiding adhesion of the retina to the RPE such as the interphotoreceptor matrix and interdigitation of rod outer segments with RPE microvilli.
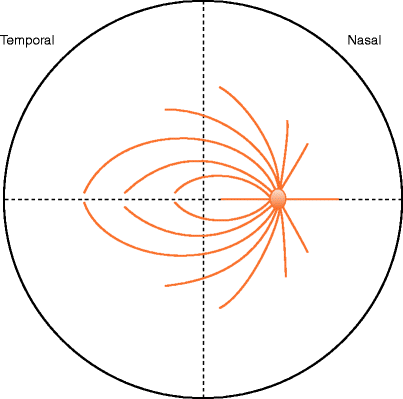
Fig. 1.7
Always consider in which direction the nerve fibres run on the surface of the retina when operating so as to minimise damage to the fibres. Try to cut or scrape along the direction of the nerve fibres rather than perpendicular to their direction
Interestingly, evidence of vitreous collagen metabolism (C-propeptide levels of type II pro-collagen) (Itakura et al. 2005) is found in the vitreous cavity of vitrectomised eyes, although hyaluronan levels are reduced.
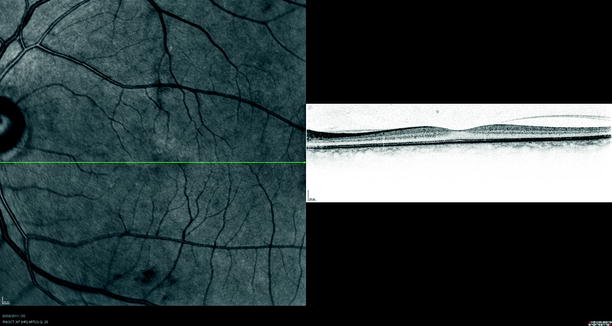
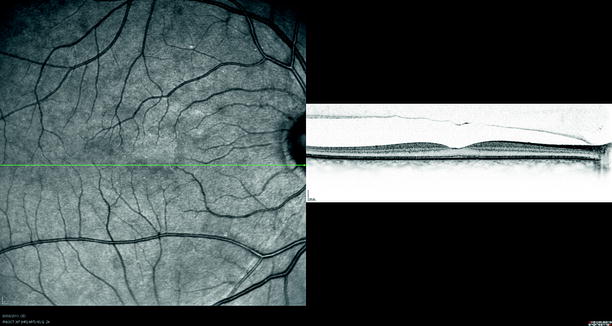
Fig. 1.8
The vitreous is more strongly attached to the fovea. The posterior hyaloid membrane can be seen detached from the temporal retina but still attached at the fovea and nasal to the fovea on optical coherence tomography (OCT)
Fig. 1.9
The vitreous is more adherent to the optic nerve head. The posterior hyaloid membrane can be seen separated from the retina but attached to the optic nerve head
1.2 Clinical Examination and Investigation
1.2.1 Using the Database
Keeping an electronic patient record of the clinical details of the patient aids clinical audit and governance. A fully operational system has been provided which runs on Microsoft Access (you will need a copy of Access 2010 to run the database) and follows the classification of disease used in this text. The structure of the database includes tables into which data is stored as a record. The main table is ‘mailing list’; the other tables are linked to this (related) to store clinical data. There are forms to input data and reports for printing data out. Learn to create queries to analyse your data. A comprehensive manual has not been provided, but with trial and error, you should soon find your way around.
1.2.2 Examination of the Eye
Examination Technique
Visual Acuity
LogMar values are recommended for the ease of analysis of data for surgical audit and governance. This can be measured by Snellen chart or EDTRS chart but requires full refractive correction.
The Slit Lamp
The vitreoretinal surgeon must be able to use the slit lamp, Goldmann tonometry and various contact lenses or three-mirror contact lens and be able to visualise the vitreous by looking behind the posterior lens. The vitreous must be inspected for clarity, cellular infiltration and by asking the patient to move the eye to inspect the mobility of the vitreous. The slit lamp allows the use of specialised lenses for the examination of the vitreous and retina, for example, super-field, 90D or 60D lenses and noncontact lenses.
Table 1.1
Various lenses and their characteristics
Lens | Slit lamp or BIO | Field of view | Magnification | Depth perception | Uses | Periphery |
|---|---|---|---|---|---|---|
20D | Bio | Good | Fair | Fair | Peripheral | Far periphery |
28D | Bio | Very good | Poor | Poor | Small pupil or paediatric | Far periphery |
90D | Slit lamp | Poor | Good | Fair | Small pupil | Posterior to the equator |
Super-field | Slit lamp | Good | Good | Fair | General | Equator |
60D | Slit lamp | Poor | Very good | Good | Macula | Not for the periphery |
Goldmann three-mirror | Slit lamp | Poor | Very good | Good | General | Equator |
Hruby lens | Slit lamp | Poor | Very good | Very good | Macula | Nil |
Rodenstock | Slit lamp | Very good | Poor | Poor | Panretinal photo coagulation | Equator |
Binocular Indirect Ophthalmoscope
A principle extra skill required is the use of the binocular indirect ophthalmoscope with indentation of peripheral retina (Schepens 1947).
Method: Examine in a systematic manner. Always lay the patient flat on an examination couch preferably with no pillow.
Note: Commence examination standing at the patient’s side whilst examining the 12 o’clock position of the retina, and move around the head of the patient to the other side of the patient systematically examining the whole of the peripheral retina returning to the 12 o’clock position.
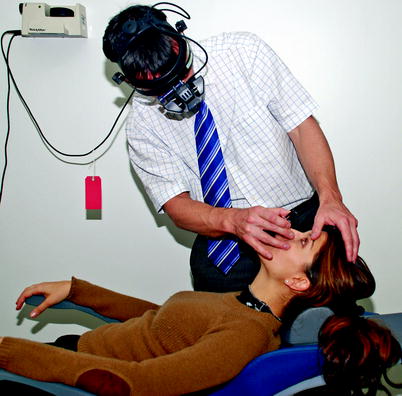
Fig. 1.10
Notice the sideways lean when examining with the indirect ophthalmoscope in order to avoid backache
Remember, the patient looks in the direction of the retina that the observer wishes to see; for example, if examining the superonasal retina, the patient looks superonasally. Initially examine the eye without indentation thereby orientating to the distribution of subretinal fluid (SRF) and to provide a clue to localisation of features such as retinal breaks according to Lincoff’s rules (described in Chap. 5). Using indentation to examine the retina, move around the patient’s head in the opposite direction finally returning to the original starting position.
Using the Indenter
The superior retina (and the temporal side) is often easier to examine because the upper lids are easier to indent through. Ask the patient to look down, place the indenter head on the lid above the tarsal plate and ask the patient to look up. As they do this, rotate the indenter superiorly and apply pressure in the globe aiming towards the centre of the eye.
Note: Get the feel for the required pressure to apply to the globe by pressing the indenter gently on your thumb just enough to depress the skin of the tip of your thumb. Try not to push back into the orbit which only succeeds in indenting the orbital septum and causing the patient discomfort.
Observe the indent on the retina whilst gliding the indenter back and forwards or from side to side. Watching the movement of the retina facilitates retinal tear detection. Aim to be able to examine the retina right up to the ora serrata; small anteriorly placed holes can be difficult to find and will cause failure of surgery if undetected. When examining the inferior retina, ask the patient to look up, place the indenter below the inferior tarsus and then ask the patient to look inferiorly. The horizontal positions are difficult to see because the canthal tendons and caruncle make indentation uncomfortable. Orientate the indenter vertically, place the indenter above or below these structures and move it sideways to move them out of the way for indentation.
Use a metal indenter, either the modified thimble variety or the pen-sized stick.
Ultrasonography
Fig. 1.11
B-scan ultrasound is essential for the running of a vitreoretinal clinic and should be present in every clinic and operated primarily by the vitreoretinal surgeon himself for interpretation of the dynamic signs in the eye
Ultrasound is essential for the examination of the eye with medial opacities (Fig. 1.11).
Note: Learn to perform this technique. Ultrasound is a dynamic examination from which information can be rapidly obtained in the clinical setting.
Ultrasound has a frequency, number of cycles/s of 20 hertz (Hz) which is inaudible to human ears. The higher the frequency of the ultrasound (the shorter the wavelength), the higher is the resolution but at the cost of less penetration into tissues.
Appropriate frequencies for ophthalmology vary from 7.5 to 10 MHz for the posterior segment and with 20–50 MHz for the anterior segment.
Sound travels faster through solids than liquids, for example, through both aqueous and vitreous at 1,500 metres/second (m/s) and through the cornea and lens at 1,650 m/s. Therefore, clear images are readily available from the eye and orbit.
Sound is reflected when it encounters an interface of different tissue densities resulting in an echo whose strength relates to the difference in the densities. The signal is highest when the interface is perpendicular to the ultrasound beam.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


