Toxoplasmosis
Khalid F. Tabbara
Toxoplasmosis is a common disease caused by the organism Toxoplasma gondii. The disease is widespread in nature and can affect both humans and animals. Toxoplasmosis can be an acquired or a congenital disease, and it is considered to be the most common cause of infection of the human retina.1,2,3,4,5 The organism affects the posterior segment of the eye, leading to relentless destruction of vital structures of the retina which may result in loss of vision.6,7,8 The disease may recur in the eye without any evidence of systemic manifestation or increase in the antibody titer. Toxoplasma affects both immunocompromised and immunocompetent individuals. Following an acute infection, the organism may become encysted in the host and persists in tissues for months or years without clinical manifestations.9,10,11 The persistence of Toxoplasma antigens in the host may assure long-term immunity in the absence of immunologic compromise. In immunocompetent individuals, the clinical manifestations are subclinical in most patients, but sometimes can cause flulike illness without serious complications. The ocular lesions, on the other hand, may lead to serious complications and loss of vision in both the immunocompetent and the immunucompromised host. The active phase of the disease is self-limited, but because of the persistence of the Toxoplasma cyst in the tissue, recrudescence of the disease can occur from unknown factors or immunologic compromise. The disease can recur in the eye without any evidence of systemic manifestations or increase in the antibody titer. Toxoplasma is an intracellular parasite, and cells in the retina are commonly affected. The vitreous, which is acellular, is rarely invaded by the organism, but Toxoplasma-infected inflammatory cells may reach the vitreous cavity. The retina sustains the major damage and primary insult. Recent increase in the human immunodeficiency virus (HIV)-infected population led to an increase in the serious ocular disease caused by Toxoplasma.12,13,14 Early diagnosis is essential for prompt initiation of therapy, especially when the lesion is close to the macula, optic nerve head, and papillomacular nerve bundle.15,16,17,18 In many cases, the toxoplasmic retinochoroiditis clinical picture is typical, but there are instances in which the disease can mimic other entities or present with unusual clinical manifestations.19,20 At times, toxoplasmosis leads either to posterior granuloma or panuveitis leading to misdiagnosis of the entity.21 Because Toxoplasma is the most common cause of posterior uveitis, ocular toxoplasmosis should always be in the differential diagnosis of diffuse panuveitis or posterior uveitis.
The Organism
Toxoplasma gondii is an obligate intracellular protozoan belonging to the phylum Protozoa, the subphylum Apicomplexa, class Sporozoasida, order Eucoccidiorida, and family Sarcocystidae. The three forms of the parasite are the tachyzoite, the bradyzoite, and the sporozoite, can cause infection in humans. The tachyzoite is a 6 μm to 8 μm in length and 2 μm to 3 μm in width (Fig. 1). The organism is motile and stains with both Giemsa and Wright stains. It is easily propagated in the peritoneal cavities of mice, as well as in mammalian tissues cell cultures. The tachyzoite has the ability to replicate in all mammalian nucleated cells. In the host cells, tachyzoite multiplies by endodyogeny. This reproductive process is susceptible to heat, freezing, thawing, desiccation, and gastric enzymes. Under electron microscopy, the organism appears to have a pointed anterior end with an apex referred to as the apical complex containing the conoid and having specialized subcellular structures. The organism has a blunt posterior end. The organism enters cells by its conoid. The tachyzoite moves by gliding, twisting, wiggling, and rotating. These movements are assisted by cytoplasmic fine freely floating filamentous structures in the cytoplasm attached to the anterior and posterior conoidal rings in the apex of the parasite. The conoid at the apical end has cellular substructures that help it expand, retract, tilt, and rotate. These are important maneuvers and such motion is required for the penetration of the cells.
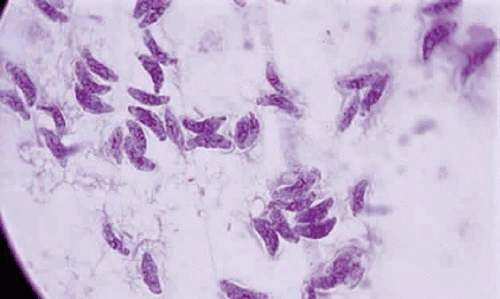 Figure 1. Tachyzoites of Toxoplasma gondii stained with Giemsa (×100). |
Following a phase of rapid proliferation, the organism starts to form pseudocysts and later true cysts. In tissue cultures, pseudocysts begin to form as early as 4 days after inoculation of the organism. Rapid encystment occurs if the environmental conditions are unfavorable.11,22,23 The bradyzoites are inside the cyst. The cyst can contain 50 to 3,000 tachyzoites and has a diameter reaching up to 250 μm. The cyst wall is strongly acidophilic and periodic acid Schiff positive. It contains constituents that are derived from both the parasite and host tissue. These structural elements are derived from the host tissue and are not recognized by the immune system, which afford protection of the parasite.
Bradyzoites inside the cyst divide at a very slow rate and are metabolically depressed. Because the tissue cyst incorporates elements derived from the host, it is easily tolerated by the host and no inflammatory reactions are seen around it. The cyst can persist in tissues, such as the central nervous system, heart muscles, or eye, without provoking any inflammatory reactions. The bradyzoites inside the cyst derive their nutrition from the slow diffusion of substances through the cyst wall. The organisms increase in number within the cyst in the retina, and once the cyst wall breaks down, the bradyzoites escape, converting into tachyzoites, which in turn invade contiguous cells. The bradyzoites inside the cyst derive their nutrition from the slow diffusion of electrolytes as well as amino acids, glucose, water, and gases. The cyst wall is a complex structure of both proteins and polysaccharides. Conversion of the tachyzoites to a bradyzoites can be noted in the infected retinal tissues of both animals and humans (Figs. 2 and 3).
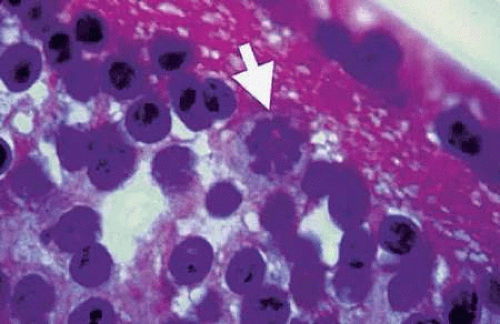 Figure 2. Bradyzoites of Toxoplasma gondii inside cysts of tissue sections of chronically infected rabbit’s retina (arrow). The animal was killed 6 months following acute Toxoplasma retinochoroiditis following suprachoroidal injection of 1000 organisms (Beverley strain). No clinical evidence of inflammation was seen in the retina when the animal was killed (H&E, ×400). |
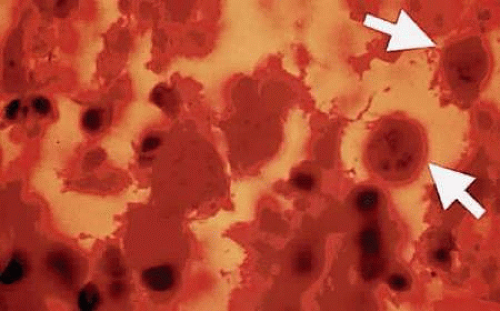 Figure 3. Toxoplasma cysts containing bradyzoites in a human retina (arrows). The eye was enucleated because of absolute glaucoma. The patient had been treated with oral and periocular corticosteroids (H&E, ×400). |
Life Cycle
Toxoplasma has sexual and asexual reproduction. The cat and its related feline species are the known definitive hosts of Toxoplasma gondii. Both domestic and certain wild cats maintain the sexual reproduction of organisms. The intermediate hosts are a wide variety of animals, including humans. In the definitive host, the parasite has both enteroepithelial and extraintestinal cycles. The enteroepithelial cycle of Toxoplasma is the sexual cycle; it leads to gametogony with oocyst production and sporogony takes place after excretion. The extraintestinal phase in cats is an asexual cycle and occurs in the tissues outside the intestine.
Three to six days following ingestion of cyst-infected meat, the bradyzoites are released, and enter in the epithelial cell lining of the small intestine particularly the jejunum. Bradyzoites undergo five sequential development stages that pass through several generations by endodyogeny, endopolygeny, splitting, and or schizogony. On reaching the final three stages, schizonts form microgametes and macrogametes in the small intestine. The two gametes form zygotes, which develop into oocysts.24 The oocyst detaches from the intestinal epithelium and is excreted in the feces. Sporulation in the oocyst occurs in 1 to 5 days, depending on temperature and availability of oxygen. Sporocysts are formed, each of which contain four sporozoites. On ingestion of the bradyzoites in the cyst of infected animal tissues, the enteroepithelial cycle in the cat is initiated, whereas ingestion of oocysts containing sporozoites can lead to generalized infection in the cat. Following oocysts excretion by the cat, sporulation occurs. The mature oocyst remains infectious in moist soil for long period of time if not subjected to extreme climatic condition. Freezing to -20°C or heating above 66°C and desiccation are lethal to the cyst. Following ingestion of the oocyst, the wall is digested by gastric enzymes releasing the sporozoites that can invade the intestinal mucosa and lead to extraintestinal systemic dissemination that infects the tissues such as the muscle, heart, brain, lung, lymphoid organs, and eye. The organism multiplies rapidly by endodyogeny, leading to disruption of the infected cell membrane, thereby liberating the tachyzoites which enter contiguous cells. Thus the rapidly proliferating tachyzoites are responsible for the initial spread of the infection and tissue destruction in the acute stage. In response to hostile environmental conditions, tachyzoites transform into slowly multiplying organism or bradyzoites that form true cysts in tissues (chronic stage of the disease). The cysts can remain in tissues throughout the life of the host, but can cause reactivation of the clinical disease when the host immunity is suppressed. Toxoplasma, therefore, can be perpetuated in acquired and congenital transmission without involving the definitive host of the parasite. The intermediate hosts would support the asexual life cycle of the parasite.
Animal Models of Toxoplasmosis
Ideal animal models of infectious diseases of the retina should have retinal lesions that morphologically and histopathologically resemble lesions seen in humans. The natural course of the disease in selected animal models should be similar to the disease seen in humans. The retinochoroiditic lesions should be readily reproducible, and the immunologic defense mechanisms of the animal must be similar to the defense mechanisms of humans. Several animal models of Toxoplasma retinochoroiditis have been described.25,26,27,28 Injection of Toxoplasma organisms (Beverley strain) into the suprachoroidal space of pigmented rabbits produced focal retinochoroiditis with no encephalitis.26 This animal model was used to study the pathogenesis and the effects of therapeutic modalities on the course of Toxoplasma retinochoroiditis. Murine Toxoplasma retinitis can be induced by intraperitoneal injection of T. gondii. Mice develop encephalitis and retinitis and succumb to the disease. Following intraperitoneal injection of Toxoplasma in mice, the disseminated infection led to Toxoplasma cysts in the retina and brain (Fig. 4).
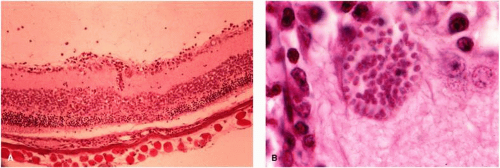 Figure 4. Toxoplasma chronically infected mouse showing retina-choroid tissue section with inflammatory reaction and Toxoplasma cyst in the inner retinal layer (arrow). Insert shows an enlarged view of the Toxoplasma cyst. |
An animal model of ocular toxoplasmosis was produced in a nonhuman primate, the cynomolgus monkey, by Culbertson et al.27 The retinochoroiditis was induced by direct inoculation of Toxoplasma organisms into the retina through the pars plana under direct microscopic visualization. The focal necrotizing retinitis lesions had morphologic and histopathologic resemblance to the lesion seen in humans, and the course of the retinochoroiditis closely resembled the course of the disease in humans. The cynomolgus monkey was found to be resistant to the RH strain of T. gondii and developed no evidence of systemic disease and no encephalitis. Furthermore, the immune mechanisms of the animal model are similar to those of humans. Healing of the retinochoroiditis was associated with scar formation, with atrophic scar with variable pigmentation. Histologic studies revealed mononuclear inflammatory reaction with evidence of areas of necrosis seen in the retina (Fig. 5). Toxoplasmic retinochoroiditis in subhuman primates appears to be a good animal model for the study of ocular toxoplasmosis.
 Figure 5. Section of the retina of a cynomolgus monkey infected with T. gondii shows extensive retinal necrosis with disruption of the normal retinal architecture, mononuclear cell infiltration, and severe choroiditis (H&E, ×100). |
Epidemiology
Modes of Transmission
Toxoplasmosis can be acquired in humans by several modes of transmission, which are outlined in Table 1. The most common modes of transmission of the disease are by ingestion of undercooked Toxoplasma cyst-containing meat and the inhalation or ingestion of oocysts. Oocysts are resistant to environmental factors of extreme temperatures.
Table 1. Modes of Transmission of Toxoplasma | |
|---|---|
|
Toxoplasma-infected animals that have developed a subclinical infection, can live normally but harbor the encysted form (bradyzoites) of the parasite in their muscular system. Undercooked beef, mutton, pork, and chicken have been incriminated in the transmission of the disease.29,30,31,32,33
Water-borne transmission of the disease may represent a major mode of transmission of the infection in many countries around the world.1,34 The presence of infected kittens around homes may allow the accumulation of great numbers of oocysts. Teutsch et al 35 have reported an outbreak of toxoplasmosis in which Toxoplasma oocysts were the source of infection and the presumed mode of transmission was by inhalation.
Accidental contamination of hands while disposing of cat litter represents another mode of transmission of the disease. Person-to-person transmission of toxoplasmosis can occur with blood transfusion and organ transplantation. An infected allograft organ that was prepared for transplantation has also been shown to transmit Toxoplasma in humans.36 The consumption of raw milk can also lead to transmission of the disease.37
If a woman acquires the infection during pregnancy, the disease can be transmitted to the fetus. This form of transplacental transmission is of great clinical importance. Toxoplasmic infection in consecutive siblings is rare, but reports of supposed congenital ocular toxoplasmosis have been described in siblings.38 The prevalence of congenital toxoplasmosis has been estimated to vary between 1:1,000 and 1:10,000. In England and Wales, the prevalence of congenital toxoplasmosis is 1.62 in 100,000.39
Most of acquired toxoplasmosis is asymptomatic. The prevalence of toxoplasmosis among adults in the United States ranges from 20% to 70%, depending on region and age. It is estimated that one-third of the world population is infected. Serologic evidence of T. gondii antibodies may vary from one population to another. The prevalence of toxoplasmosis is influenced by age, geographic location, and social and economic factors.32 Individuals living at high elevations (above 5,000 feet) appear to have a lower rate of infection than matched controls at sea level. The disease frequently occurs in hot and arid areas as well as in tropical areas.30,31,32 The oocysts appear to resist environmental conditions. Populations with increase in their consumption of undercooked meat may be more affected that other population. In western Europe and areas of Central and South America where the individuals consume undercooked meat, the rates of antibodies to T. gondii may exceed 90% by the fifth decade.40 The presence of Toxoplasma antibodies increases with age; the older the individual, the greater the prevalence of antibodies to T. gondii. Ocular toxoplasmosis may be associated with acquired systemic toxoplasmosis or may occur months or years after an episode of systemic toxoplasmosis. It is estimated that 25% of pork and mutton contain Toxoplasma cysts.29 Toxoplasma has also been isolated from egg and milk. The cooking habits and hygiene may influence the transmission of toxoplasmosis.
In the Netherlands, positive antibodies to T. gondii in the general population between the ages of 25 and 45 years is 57%. In central Europe, the prevalence of Toxoplasma antibodies ranged between 42% and 56%. The highest prevalence of Toxoplasma antibodies among pregnant woman was found in Brazil, ranging from 59% to 78%.40
Burnett et al.1 reported the largest outbreak of acquired toxoplasmic retinitis in 20 patients. Of 20 patients, 22 eyes developed active toxoplasmic retinochoroiditis following systemic disease.1 Patients infected with HIV had significantly higher prevalence of Toxoplasma antibodies than apparently healthy individuals.41
Transplacental transmission of the disease and congenital toxoplasmosis can occur in mothers who acquired the infection during their pregnancy. The risk of severe clinical disease in the fetus occurs in the first trimester of pregnancy and the severity of the clinical disease decreases with the increase of the fetal age. Children infected with congenital toxoplasmosis in the first trimester of pregnancy and who had normal ultrasound findings were studied by Berrebi et al.42 Of 36 infected children, 28 (78%) presented with subclinical congenital toxoplasmosis. Seven (19%) children developed retinochoroiditis with major vision loss. All children had normal intellectual development and one child (3%) developed severe congenital toxoplasmosis. Because 97% of infected children with congenital toxoplasmosis during the first trimester of pregnancy were asymptomatic, the authors recommend that pregnancy in mothers who acquire toxoplasmosis in the first trimester should not have a therapeutic abortion.42 In England and Wales, Gilbert39 found low incidence of symptomatic congenital toxoplasmosis. Their findings confirmed and supported the current policy not to offer prenatal or neonatal screening for Toxoplasma infection. On the other hand, it is highly recommended to establish strategies for the prevention of acquired infection during pregnancy and in childhood. Among 38 children with confirmed Toxoplasma infection, 22 (58%) were classified as having congenital toxoplasmosis.39 Of those, 10 (45%) had retinochoroiditis. It is estimated that 50% of children with congenital toxoplasmosis will develop retinochoroiditis. Early treatment of congenital toxoplasmosis can prevent the sequelae of the disease.39
Immunosuppression may lead to recurrences of toxoplasmic retinochoroiditis. Immunosuppression can be induced by debilitated illness, the use of immmunosuppresive agents or malignant disorders such as hairy cell leukemia and Hodgkin’s lymphoma.43,44 Pregnant women who have retinochoroiditic scars may develop recurrence of retinochoroiditis during their pregnancy.45,46 Bosch-Driessenet al.47 have shown that reactivation of ocular toxoplasmosis occurs within 4 months following cataract extraction in 36% of the patients. In their study, other risk factors for the development of recurrences of ocular toxoplasmosis were identified. Patients who had phacoemulsification have a tendency to have a decrease in the reactivation of toxoplasmosis, presumably because of minimized surgical trauma. It is also suggested that ultrasonic waves may induce rupture of the cyst.47 The effects of ultrasonic waves on toxoplasmosis in retinal tissues have not been studied. The finding of reactivation of toxoplasmic retinochoroiditis following cataract surgery may justify the chemoprophylaxis with antimicrobial agents in patients who have healed retinochoroiditic scars and who are having cataract surgery. Stress and chronic anxiety can also lead to reactivation of toxoplasmic retinochoroiditis in patients who have retinochoroiditic scars. Examples are college students undergoing examination. The stress of the examination may lead to reactivation of toxoplasmic retinochoroiditis. Infection with HIV may also lead to immunosuppression and reactivation of toxoplasmic retinochoroiditis. Furthermore, it was observed that patients who are in the second and third decade of life may tend to have a higher rate of recurrences of toxoplasmic retinochoroiditis than patients who are in the older age group with retinochoroiditic scars.
The risk factors for transmission of T. gondii in children and adults are predominantly owing to ingestion of undercooked meat, including mutton, pork, beef, and chicken. Inhalation of the oocysts containing the sporozoites can also lead to toxoplasmosis.
Pathogenesis
Intracellular Parasite Invasion
Because the protozoan cannot survive in the extracellular space, the initial step of entry into the host cells is a critical maneuver for the survival of the organism. The tachyzoite has a well-defined nucleus that is oval or round with a central karyosome. Toxoplasma utilizes various strategies to invade host cells. In general, intracellular parasites tend to subvert cellular signaling pathways to gain a foothold against host defenses. Most intracellular parasites gain entry via host mediated mechanisms, but the apicomplexan organisms, such as Toxoplasma, use a system of adhesion-based motility called gliding to actively penetrate the host cell. Motility of the organism depends on the active polymerization between the filamentous intracellular structures facilitating the migration of the parasite to cross the cell membrane barriers and to invade the host cell. This efficient invasion utilizing unique strategies makes the apicomplexan pathogens dangerous. Certain enzymes, such as polymorphic secreted kinases, are key for the virulence factors in toxoplasmosis.51,52 Furthermore, the Toxoplasma MIC2 protein represents a major determinant of invasion and virulence of the parasite.53 The parasite has a conoid rostral end with an ability to thrust outward at the time of invasion into the cell. At the time of contact between toxoplasmic plasmalemma and the host cell membrane, the differentiated organelles in the conoid, known as rhoptries, secrete their enzymatic contents, which lead to disruption of the host cell membrane, allowing the organism to get access to the cytoplasm. This process of active penetration takes 34 to 305 seconds under laboratory conditions54 and the parasitophorous vacuole is formed from the host cell plasma membrane. The thrusting of the conoid into the cell membrane is probably accomplished by an actinlike system that helps in the erection and rigidity of the conoid. The organisms enter the retinal host cells by active invasion and enter the macrophage by both phagocytosis and active invasion. In naïve macrophages, the organism has the ability to actively invade the macrophage, which would provide protozoan with a free ride that helps in the dissemination of the disease. In immunized hosts, the organism attains the intracellular status in the macrophage by active phagocytosis. The parasite may enter the retinal pigment epithelial cell by active penetration or by phagocytosis. In most host cells, however, invasion appears to be the predominant means of cellular entry of the organism. The difference between invasion and phagocytosis is recognized by the presence of microfilamentous aggregates in the conoid by electron microscopy. Several strategies are adopted by Toxoplasma for its intracellular invasion.55 Contact between the macrophage and the anterior tip (conoid of the parasite) may augment the penetration of the protozoan into the macrophage. If a macrophage grasps the posterior end of the organism, rhoptry discharge may not occur. The absence of filament aggregates under the cell membrane of the macrophage during toxoplasmic penetration is strong evidence that the cellular invasion was a parasitic affected process. Several factors may interfere with cellular penetration, whether the penetration is by phagocytosis or by active invasion. Coating the organism with IgG or C3 stimulates and enhances phagocytosis. In the absence of serum, phagocytosis may proceed, but very slowly. Active invasion by the parasite may also be inhibited by cytochalasin D, a substance that interferes with actin filament function.56 This suggests that the conoid filamentous structure is not only a membrane but also is supported by an actin-myosin system that helps the organism to thrust forward and move in an undulating motion.
When the parasite attains intracellular habitat, it starts to multiply, leading to host cell destruction. Live organisms do not stimulate a respiratory burst during phagocytosis by the macrophage.57 The parasite may retain some of the disrupted macrophage plasmalemma adherent to the surface immediately after invasion. A vacuole quickly forms around the parasite that has invaded the cell. It appears to be larger than the organism with tubules connecting the limiting membrane with the plasmalemma of the parasite.58 Each vacuole contains one parasite, and sometimes several organisms may be seen within a single macrophage. In activated macrophages, however, the organism will have difficulty surviving within the macrophage, and the parasite is killed following fusion between the cell membrane and the internal limiting membrane of the vacuole causing the microbiocidal activity. The organism, in general, is resistant to hydrogen peroxide, but it is susceptible to oxygen intermediates generated by the xanthine oxidase system.57
Toxoplasma gondii invades mammalian cell by using its unique form of gliding motility. Recently, host cell membrane adhesion molecules were identified. These molecules are capable of adhering to external receptors, suggesting that the parasite has multiple options for host cell recognition and invasion. The host cell membrane adhesion MIC2, in consort with its partner protein M2AP, participates in the invasion pathway. Reduced MIC2 expression resulted in confusion of the organism and mistrafficking of M2AP leading to a defective host cell interaction attachment and invasion.53 The loss of gliding motility and the inability to support the lethal infection in the host cell was caused by inhibition of MIC2 expression. Mice infected with organisms that are MIC2 deficient had significantly lower parasite burden in infected tissue and attenuated the inflammatory immune response as well as induction of long-term protective immunity. These experimental findings if borne in mind will make this unique way to approach a potential vaccine for toxoplasmosis. The findings demonstrated that MIC2 protein complex is a major virulence determinant of T. gondii and that MIC2-deficient parasites constitute an effective live attenuated vaccine for experimental toxoplasmosis.53 Macrophages in the intestinal wall are stimulated by the abundance of lipopolysaccharides available in the mucosa from gram-negative bacteria59 T. gondii has the ability to block the response of macrophages to lipopolysaccharides, possibly avoiding the macrophage killing response. The MIC2 and M2AP are released from the micronemes of the parasite as well as the rhoptries. T. gondii appears to play a major role in the recruitment, maturation, and migration of human dendritic cells derived from CD34+ progenital cells. The action of the parasite is promoted by the release of soluble factors. Soluble factors derived from type I and type II T. gondii strains appear to possess a chemokinelike activity that attracted immature dendritic cells.60 Host cell invasion is an essential step during infection by T. gondii. Recent evidence strongly suggests that protein charged and released from the apical conoid of Toxoplasma secretory organelles (micronemes, dense granules, and rhoptries) play a key role in host cell infection and survival during infection. It appears that T. gondii exploits host enteroepithelial macrophages to prevent lipopolysaccharide triggered interleukin 12 (IL-12) and tumor necrosis factor-α (TNF-α) productions. These suppressive effects released by the parasite have an immense effect on macrophage cytokine production, which allows the organism to survive in a less hostile environment.61
Virulence
Pathogenesis of toxoplasmosis depends on the virulence of the Toxoplasma organism’s ability to establish a chronic disease, and the immune factors of the host. Although it has a sexual phase of the life cycle, the disease is caused by the T. gondii organisms via its asexual reproduction and transmission from other intermediate hosts. T. gondii has three clonal lineages that predominate in North America and Europe.62 The three clonal lineages are referred to as types I, II, and III. The widespread of these clonal lineages is attributed to the circumvention of the sexual cycle by the new mode of transmission via the asexual transmission. Type I clonal lineage is highly virulent to humans and animals.63,64,65,66 It has been shown that type I lineage was detected by multiplex vested polymerase chain reaction (PCR) analysis in cerebral spinal fluid of 8 of 10 patients who were HIV positive.64 Lineage types II and III are less virulent than type I. Examples of type I strain of the T. gondii is the RH strain and type II is the ME49. Su et al.66 have shown that the acute virulence of T. gondii is regulated by a link to SAG1 locus on chromosome VIII in T. gondii. In experimental animals, the acute virulence strain of T. gondii is type I, which is uniformly lethal to mice. Following intraperitoneal injection of the RH strain of T. gondii in mice, the animals develop encephalitis and die within 4 to 6 days after the injection. Analysis of naturally occurring recombinant strains of T. gondii that carry allele 1 at the SAG1 locus is characteristic of type I strains of the organism and has been previously correlated with the acute virulence phenotype of Toxoplasma.66
Production of IL-12 by macrophages infected with T. gondii depends on the parasite genotype.67 Infection with the type II strain of T. gondii stimulated the production of IL-12. On the other hand, type I strain, which is the virulent strain of T. gondii, does not stimulate IL-12 production except after active invasion of live parasite. For this reason, the findings suggest that early induction of high levels of IL-12 by macrophages infected with type II strain parasites may contribute to more effective control than infections with type I clonal lineage. The conversion of tachyzoite to bradyzoite in an encysted form takes place when the environmental conditions are not favorable for the organisms. Arginine deprivation efficiently blocks the replication of intracellular T. gondii. Arginine starvation was found to trigger efficiently the differentiation of tachyzoites into bradyzoites contained within stable cyst-like structure.68 The same parasites expressing bradyzoites antigen can be efficiently switched back to proliferating tachyzoites several weeks after arginine starvation. Other factors may play a role in the cyst formation of the parasite to be able to survive and to secure propagation of the phylogeny. Transformation of the tachyzoite stage into the latent bradyzoite cyst underlies the chronic disease and leads to a lifetime risk of recurrence of toxoplasmic retinochoroiditis.
Immune Response to Toxoplasma
Acute systemic disseminated Toxoplasma infection usually occurs both in immunologically normal and immunocompromised hosts. Most systemic infections in normal hosts are subclinical. The disease is brought under control by the normal cellular and humoral immunity that develops following the acute infection.
The parasite reaches the ocular structures via the bloodstream and establishes a focus of infection in the ocular compromised tissue, the retina. The protozoan enters retinal cells and establishes a focus of retinitis that soon progresses to involve the choroid. In vivo, the focus of retinitis is characterized by infiltration with mononuclear cells. The Toxoplasma cyst remains initially inside the cell and, with time, the host cell membrane gets disintegrated and the cyst remains in tissues and adjacent to an inactive retinochoroiditic scar for many months or years. The bradyzoite keeps dividing at an extremely slow rate, and this leads to stretching of the cyst wall. Ultimately there is disruption of the cyst wall and the organisms escape from the cysts into the tissues. If the organisms are not controlled by cell-mediated immune mechanisms, the parasite will invade neighboring cells, causing recurrence of the infection. Reactivation of toxoplasmic retinochoroiditis may be induced by immunologic suppression states, including stress, anxiety, or other factors of immune suppression, malignancy, use of immunosuppressive agents, and acquired immunodeficiency syndrome (AIDS) (Table 2).
Table 2. Recurrences of Toxoplasmic Retinochoroiditis: Risk Factors | ||
|---|---|---|
|
The exact cause of cyst rupture has never been determined. The fluctuation of antibody titers in normal subjects with positive titer suggests periodic rupture of the cyst wall. The recurrent retinochoroiditis may represent an active infection in a predominant manner, but to a lesser extent there is a hypersensitivity response to Toxoplasma antigens and possible hypersensitivity to retinal antigens. Injection of Toxoplasma antigens into the carotid artery of immunized nonhuman primates did not cause recurrences of healed retinochoroiditic scars.69 Furthermore, immunized eyes with previous Toxoplasma infection showed no evidence of recurrence of retinochoroiditis when the Toxoplasma antigens were injected intraocularly.
Clinical observations, however, have shown that the granulomas of Toxoplasma retinochoriditis are neither self-limited nor short-lived and many progress into large necrotizing lesions. It is conceded, therefore, that the initial lesion of Toxoplasma retinochoroiditis involves active parasitic infection and multiplication of the organism within the retinal tissue. In recurrent retinochoroiditis, on the other hand, there is release of viable parasites, which actively multiply in the retinal tissues and cause reactivation of the disease. The inflammatory response may protect the tissues from the ravages of the parasite, but at the same time this may lead to destruction of the innocent bystander tissue. Following necrosis of the retinal and choroidal tissue, arteriovenous anastomosis and shunt, as well as subretinal choroidal neovascularization, may occur.
In patients with severe immunosuppression, the Toxoplasma organism may invade the tissue and lead to anterior uveitis. The Toxoplasma antigens can be detected from both the vitreous and the aqueous humor during the acute phases of retinochoroiditis.70 The anterior uveitis is self-limited once the posterior uveitis is brought under control. The inflammatory reaction in the anterior uvea may be controlled by the topical use of steroids. Increase in intraocular pressure may occur. In experimental animals and humans, retinochoroiditis is associated with a hypersensitivity vasculitis that can manifest as arteritis or phlebitis.71
Activation of natural killer (NK) cells is regarded as one of the first events to occur following infection with T. gondii. This form of resistance is innate. The NK cells have the ability to produce interferon-γ (IFN-γ) early in the course of infection, leading to a resistance to infection with T. gondii.72 This is followed by the activation of macrophages that are stimulated by both live and dead tachyzoites of T. gondii. Macrophages produce high levels of IL-12 which, in turn, will stimulate the NK cells to produce more of IFN-γ. Treatment with anti-IL-12 antibody resulted in early mortality of mice following infection.73 The activation of NK cells may be induced by both live and dead parasites. The parasites glycoprotein can stimulate the host macrophage production of cytokines such as IL-1, IL-12, and TNF-α.74 The ability of IFN-γ to stimulate NK cells depends on the presence of IL-12, TNF-α, and IL-1.75,76 The CD28-B7 interaction may play a role in the regulation of NK-cell response during infection with T. gondii. Shortly after the activation of NK cells during the course of infection, there is a short decrease in the NK cell activity, which is induced by the release of IL-10 and tumor growth factor-β (TGF-β). The production of INF-γ and IL-12 trigger the release of CD4+ cells which, in turn, will produce IL-2.77 The expansion of CD8 cells is in conjunction with macrophage cytokine-derived IL-15. IL-12 has also been shown to promote type 1 cytolytic CD8+ T cell development. Depletion of CD4+ cell population during the HIV infection may lead to dissemination of toxoplasmosis. The marked decrease in CD8+ lymphocytes may lead to reactivation of latent infection.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


