(1)
University of Sydney, Sydney, Australia
Overview (Fig. 15.1)
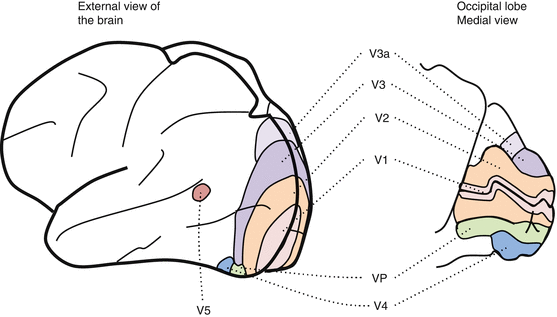
Fig. 15.1
The extrastriate cortex
The extrastriate cortex is involved in the analysis of specific attributes of visual stimuli
(e.g., color, form, movement, and binocular disparity).
Visual information is progressively decomposed as it is channeled through processing streams.
Nonhuman diurnal primates such as macaque monkeys have approximately 25 cortical visual association areas [1]; humans probably have a similar number [2].
In all primates, primary visual cortex (area V1, striate cortex, Brodmann’s area 17) occupies ~12–18% of the neocortex. Although in all primates (including humans) each extrastriate area is substantially smaller than area V1, together, they occupy ~ 25–30% of the neocortices [3]. Thus, V1 and extrastriate cortices together occupy ~ 30–40% of primate neocortices [3].
Neurons in each extrastriate area have a degree of functional specificity relating to particular stimulus attributes [4].
The main areas are V2, V3, V4, and the middle temporal (MT) area, also known as V5.
Visual inputs to extrastriate cortices originate mainly or almost exclusively in area V1.
V1 neurons projecting to a given extrastriate area tend to exhibit specific receptive field properties (e.g., direction selectivity) characterizing neurons in the extrastriate area to which they project [8].
Some extrastriate cortical areas (e.g., area MT) receive substantial direct input from the retino-recipient dorsal thalamic nuclei such as the lateral geniculate nucleus (LGN) and the retino-recipient part of the pulvinar.
Laminae of the LGN (K layers) and regions of pulvinar which project to the extrastriate cortices receive direct input from the superficial, retino-recipient layers of the superior colliculus (SC).
The Ventral and Dorsal Streams (Pathways) (Fig. 15.2 and Table 15.1)
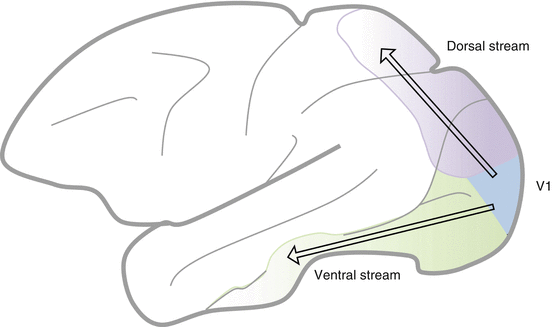
Fig. 15.2
The dorsal and ventral streams
Two broad extrastriate visual-processing streams exist:
V2 (Table 15.2 and Fig. 15.3)
Stream | Extrastriate area | Function |
|---|---|---|
V2 | Thin dark stripes: color processing Thick dark stripes: orientation selectivity and binocular disparity Pale stripes: form processing and object recognition | |
Dorsal | V5/MT V3 Parietal areas | Direction of movement, binocular disparity Dorsal V3: direction of movement Ventral V3: color, orientation Visuospatial perception and movement planning |
Ventral | V4 Inferotemporal areas | Color sensitivity, object recognition Complex receptive field properties, e.g., face recognition |
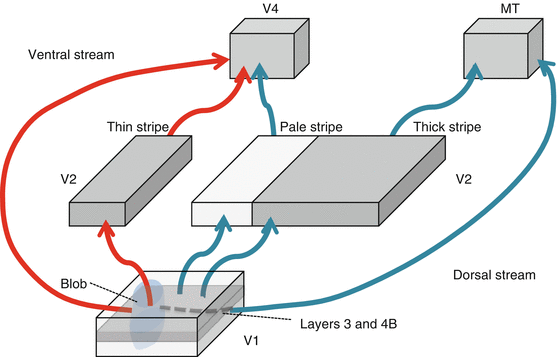
V2 receives the bulk of V1 cortico-cortical projections.
Receptive fields of area V2 neurons are 2–3 times larger than those of V1 neurons at the corresponding positions in the visual fields [16].
V2 organizes visual information for output to subsequent extrastriate processing areas [16].
It is arranged into alternating thin and thick dark stripes [17] and pale stripes based on the intensity of staining for the mitochondrial enzyme cytochrome oxidase (CO). Neurons located in the thick and thin stripes contain much more CO than neurons located in pale stripes.
Distinct visuotopic maps exist for each of the three stripe types [15].
Each stripe type represents a parallel processing pathway for stimulus attributes.
This segregation is not absolute; each stripe contains cells sensitive to a variety of stimuli [18, 19, 28].
This functional overlap probably represents integration of visual stimuli in visual processing [28, 29].
1.
V2 inputs
Input to all stripe types is from the same V1 layers and is segregated according to two pathways [30]:
(a)
CO blobs to thin stripes
(b)
Interblob areas to pale and thick stripes
2.
V2 projections
This is a ventral stream area involved in form processing, color, and object recognition [18, 19, 32].
The thick stripes project to V5, a dorsal stream area associated with motion processing [17, 18, 22].
Some thick stripe neurons project to the SC. Those projections are involved in the control of saccadic eye movement [33].
The Dorsal Stream
1.
V5/MT
2.
3.
Parietal lobe areas
The dorsal stream of visual processing terminates in the parietal lobe [48].
These projections are important for:
(a)
Constructing a spatial representation of the external world
Parietal lobe area neurons have large receptive fields that send inputs to the:
The Ventral Stream
1.
V4
V4 is located between ventral V3 and MT [48].
The V4 dorsal and ventral areas represent the lower and upper visual quadrants, respectively [48].
(i)
(ii)
V4 projections
V4 projects to inferotemporal areas involved in detailed object form analysis [55].
(iii)
V4 receptive field properties
Receptive fields of V4 neurons are substantially larger than those of V2 and V3 neurons at corresponding visual field locations.
V4 is involved in form processing crucial for object recognition [58].
Cells in V4 are principally concerned with color sensitivity; however, cells are also selective for orientation, size, and binocular disparity involved in form and shape perception [24, 59, 60].
Visual attention modulates processing in V4 [25].
2.
Inferotemporal cortex
The inferotemporal cortex receives input from V2, V4, the prefrontal cortex, and limbic system [62–65].
These cells have large, complex receptive fields selective for particular combinations of orientation, size, texture, and color [26, 66, 67].

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


