The Epstein-Barr Virus in Ocular Disease
Stephen C. Pflugfelder
James Chodosh
Epstein-Barr virus (EBV) is a human herpesvirus belonging to the subfamily Gammaherpesvirinae. EBV is a ubiquitous virus which infects the majority of humans by early adulthood.1 Epstein and associates reported the initial observation of EBV in cell lines established from biopsies of Burkitt’s lymphomas, a neoplasia which occurs predominantly in children in certain tropical regions of Africa.2 EBV was subsequently found by Henle and colleagues to be the agent responsible for infectious mononucleosis (IM), a common febrile illness.3 In addition to being associated with Burkitt’s lymphoma, EBV has been detected in other human neoplasias including nasopharyngeal carcinoma,4 thymic carcinoma,5 Hodgkin’s disease,6 oral hairy leukoplakia,7 and B-cell lymphoproliferation in allograft recipients receiving immunosuppressive therapy.8,9 The association of EBV with these neoplasias has led to the speculation that EBV is involved in the pathogenesis of these diseases. There is increasing evidence that EBV is capable of infecting ocular and adnexal tissues and causing ocular disease. The current concepts regarding the molecular genetics of the EBV life cycle, diagnosis of EBV infection, and the role of EBV in ocular disease is reviewed in this chapter.
EPSTEIN-BARR VIRUS INFECTION IN HUMANS
Primary EBV infection may manifest as IM, a disease characterized by fever, severe fatigue, pharyngitis, lymphadenopathy, hepatosplenomegaly, and occasionally hepatitis or lymphoproliferative disease, or more commonly as a mild upper respiratory tract infection which is frequently unrecognized as a primary EBV infection.1,10
The fate of EBV following infection of human cells depends on the particular type of cell infected and the pattern of gene transcription used by the virus.11 There are two major consequences of EBV infection of B lymphocytes depending on whether the active or passive latent transcriptional programs are activated. If the active latent transcription program is utilized, B cells are growthtransformed into lymphoblasts.12 EBV-infected lymphoblasts, many of which are in mitosis, can be readily detected in the blood and lymphoid tissues of patients with acute IM.13 A potent T-cell-mediated immunologic response is rapidly mounted against these virus-infected B cells, and they are cleared from the blood within several weeks. The proliferative potential of EBV is limited in vivo by HLA class I (and occasionally class II) EBV-specific cytotoxic T lymphocytes (CTLs) for the remainder of an EBV-infected individual’s lifetime.14,15 Systematic surveys identifying the immunodominant EBV epitopes suggest that most EBV-specific CTL responses are directed toward antigens expressed during active latent infection, with EBV nuclear antigens 3, 4, and 6 (EBNA-3, -4, and -6) being the most frequently identified. CTLs recognizing EBNA-2, EBNA-5, LMP-1 and LMP-2 have also been demonstrated, but at a much lower frequency and with fewer HLA class I restrictions.16 Suppression of cellular immunity in EBV-seropositive individuals (e.g., high-dose immunosuppressive therapy in transplant recipients) results in increased virus shedding in the saliva,17,18 and can lead to lymphoproliferative disease as a result of B lymphocytes expressing active latent genes that escape T-cell-mediated destruction.8,19,20
Use of the other transcriptional program in EBV-infected B cells results in “passive” latent infection. It is typically used by the virus after infecting small resting B cells, and gives rise to mRNA that encodes only EBNA-1, a nonimmunogenic protein that fails to induce an EBV-specific CTL response.16 Thus cells expressing only EBNA-1 escape T-cell-mediated destruction, and this passive latent state may represent an evolutionary strategy by the virus for lifelong persistence in immunocompetent hosts. In contrast to B cells, EBV infection of epithelial cells is a more complicated issue. Depending on the type of epithelial cell infected and the state of cellular differentiation, either a latent persistent infection may ensue or a lytic infection may occur with release of infectious viral particles.21,22,23
Similar to other herpesviruses, EBV usually persists in infected hosts following primary infection in a latent, nonpathogenic state.23,24 A number of sites of EBV persistence have been identified. Thus far, all of these sites are components of the mucosal associated lymphoid tissue (MALT) and include the major and minor salivary glands,25,26 oropharyngeal epithelia,21 epithelia in the male and female genital tracts,27,28 lacrimal gland,29 and corneal and conjunctival epithelia.30,31 At the present time, there are limited data regarding the cell type(s) in which persistent mucosal EBV infection occurs, and the repertoire of EBV proteins expressed in these latently infected cells.23,24 Because EBV DNA17 and infectious virus32 can be detected in saliva specimens of a small percentage (15% to 20%) of EBV-seropositive individuals, it appears that EBV may occasionally undergo spontaneous lytic transformation with production of infectious virus by latently infected cells in oropharyngeal MALT.
DIAGNOSIS OF EBV INFECTION
The standard for diagnosis of IM at the present time is serology. A characteristic and specific temporal pattern of appearance of serum antibodies to EBV is observed in patients with IM.10,33 By the time clinical symptoms of IM have developed, both IgM and IgG antibodies to viral capsid antigens (VCA) have reached their peak. After recovery from IM, viral capsid IgM antibodies disappear; however, anti-VCA IgG persists at low levels throughout life. Antibodies to early antigens which are indicative of viral replication, also rise during the acute phase of the disease and subsequently decrease to low or nondetectable levels in most individuals after the disease resolves. Antibodies to EBV nuclear antigens (EBNAs) appear weeks to months after the onset of IM, persist for life, and provide serologic evidence of past EBV infection.10,33,34
Additional laboratory tests may be helpful in confirming a diagnosis of IM. By the second week, patients typically have a relative or absolute lymphocytosis, and an increased percentage of atypical lymphocytes (10% to 25%).10,33 Detection of serum heterophil antibodies of the Paul-Bunnell type (Monospot test) is specific for IM due to EBV.10 These IgM antibodies, which agglutinate sheep and horse red blood cells (RBCs) and lyse beef RBCs, usually appear by the end of the first week of the infection.10 At least 10% of young adults with a typical serologic picture of acute EBV infection do not produce heterophil antibodies.35
Chronic elevations of EBV serum antibodies have been observed in a number of neoplasic and immunologic diseases. Henle and associates have reported that these serologic findings may reflect an enhanced viral carrier state manifesting as increased shedding of EBV into the saliva.36 Specific patterns of persistently elevated antibodies to EBV antigens have been found in the following EBV-associated diseases: Burkitt’s lymphoma (VCA IgG, early antigen-restricted component, or EA-R), nasopharyngeal carcinoma (VCA IgG and IgA, early antigen-diffuse component, or EA-D), recurrent parotitis in children (VCA IgG and IgA, EA-R), and chronic fatigue syndrome (VCA IgG and early antigens).37,38 Elevated EBV VCA IgG and anti-early antigen titers may also be observed in diseases with altered immunoregulation including sarcoidosis, systemic lupus erythematosis, rheumatoid arthritis, Sjögren’s syndrome, HIV infection, and immunosuppressed allograft recipients.36,39,40 The majority of reports describing an etiologic role for EBV in ocular inflammatory diseases have relied on diagnosis of EBV infection by serologic tests indicating acute or chronic infection. To conclusively prove that EBV is the causative agent for ocular diseases requires either culture of virus from or demonstration of viral proteins or nucleic acids in involved tissues.
Diagnosis of EBV infection can be made by immortalization of B lymphocytes after exposure to EBV-containing clinical specimens.1 Umbilical cord lymphocytes are the most commonly used source of B lymphocytes for EBV cell culture. Unfortunately, these cells are not always readily available, and detection of EBV-infected lymphoblasts in culture typically requires 4 to 6 weeks after inoculation.
Molecular biology techniques can also be used for detecting virus-specific proteins (antigens) and nucleic acids (DNA, RNA) in human fluid and tissue specimens. Antigens can be detected in pathologic specimens using monoclonal antibodies against EBV-specific proteins. A number of monoclonal antibodies reactive to EBV latent and lytic antigens have been developed. These antibodies have been used to detect EBV antigens in desquamated oropharyngeal epithelial cells from patients with IM,21 as well as in EBV-induced neoplasia.8,9
EBV nucleic acid sequences can be detected in specimens by hybridization with labeled nucleic acid probes specific to EBV.1 Hybridization can be performed as an in situ technique on sections cut from biopsied tissue, or on membranes onto which nucleic acids extracted from fluid or tissue specimens have been blotted.25 These techniques often lack the sensitivity for detecting the small copy number of viral genomes which may be present in the minute quantities of tissue or fluids obtained from ocular diagnostic procedures.41 The polymerase chain reaction (PCR) is a recently described molecular technique which can amplify genomic sequences from an infectious agent over one million times in several hours.42 The proven advantages of this technique for diagnosis of viral infection are its specificity for amplification of genomic sequences of a particular virus, its rapidity, and its sensitivity.42 This technique has recently been found to be sensitive enough to amplify EBV genomic sequences in tears of primary Sjögren’s syndrome patients43 and cytomegalovirus (CMV) sequences in aqueous humor specimens from patients with CMV retinitis.41 This technique hasgreat potential for evaluating the role of EBV in ocular inflammatory diseases.
MOLECULAR GENETICS OF THE EPSTEIN-BARR VIRUS LIFE CYCLE
EBV has many morphologic similarities to other herpesviruses including: (1) a protein sheet that is surrounded by DNA, (2) a nucleocapsid, (3) a protein sheet that separates the nucleocapsid and the envelope, and (4) an outer envelope composed of glycoprotein spikes, predominantly the gp350/220 protein.1 During the life cycle of herpesviruses such as EBV, a sequential pattern of gene transcription occurs as the virus progresses from the latent to the lytic phase of its life cycle.44
The entire EBV genome has been cloned and sequenced.45,46,47 Within virus particles, the EBV genome is linear and contains terminal repeats on either end of the genome (putatively thought to be important in circularization of the virus and/or integration of the EBV genome into cellular DNA), four internal repeat (IR1-IR4) regions, and five unique (U1-U5) sequence domains (Fig. 1). Approximately 100 open reading frames have been identified within the EBV genome that may potentially be transcribed and translated into proteins that are essential for the maintenance of the different phases of the EBV life cycle.44 Molecular biology techniques have allowed for the manipulation of many of these EBV genetic sequences. As a result, these in vitro studies have provided insight into the potential function of many of the EBV genes and how infection may be controlled.
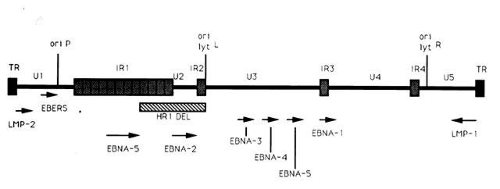 Fig. 1. Diagram of the EBV genome indicating the terminal (TR) and internal (IR) repeat regions, unique (U) regions, and the location of latent genes, several of which (EBNA-2 and LMP) may be essential for growth transformation of B lymphocytes. Arrows indicate length and direction of transcription of latency associated mRNAs. The origin of DNA replication in latent (ori P) or lytically (ori lyt, right and left) infected cells is noted. |
Although it is known that adsorption of EBV to a permissive cell occurs by interaction of the EBV gp350/220 envelope glycoprotein with the EBV/complement receptor, CD 21, on the surface of the cell,48 it is not clear how the envelope or nucleocapsid is dissolved or how the EBV DNA penetrates the cell nucleus. It is known that once the EBV genome enters the nucleus, it circularizes and that the cellular phenotype is a key factor in determining the pattern of EBV gene transcription and the consequences of virus infection on the cell.49
EXPRESSION OF EBV GENES DURING THE LATENT STAGES OF INFECTION
Two types of latent EBV infection have been identified: (1) active latent infection with transcription of EBV oncoproteins that transform (or immortalize) B cells into lymphoblasts,12 and (2) passive latent infection that is typically utilized by the virus after infecting small resting B cells and during which transcription of only one gene (EBNA-1) has been identified.
A limited number of EBV genes have been reported to be expressed during the active latent (or growth transformation) phase of EBV infection (see Fig. 1).44 These include (1) six EBV nuclear antigens (EBNA-1, -2, -3, -4, -5, -6); (2) the latent membrane proteins (LMP-1 and LMP-2); and (3) two small, nonpolyadenylated RNAs (designated EBER-1 and EBER-2).11 mRNAs encoding each of the EBNA proteins are generated by individual splicing of long rightward transcripts initiated at one of two promoters, one located in the Bam HI C region within the U1 region of the genome, and the other in the IR1 region of the genome. The LMP transcripts are expressed from separate promoters which run in opposite directions, but share the same bidirectional control region.
The EBNA-2 protein appears to be important in the transformation of B lymphocytes into immortalized cells because mutant EBV strains lacking this gene (such as the HR-1 strain with the deletion noted in Fig. 1) are incapable of immortalizing B lymphocytes.44 One of the mechanisms by which EBNA-2 may modulate growth transformation is through its ability to upregulate latent membrane protein-1 (LMP-1) transcription.50
LMP-1 is one of the most abundantly transcribed genes in EBV infected lymphoblastoid cell lines (LCLs).51 LMP-1 has recently been reported to be essential for B-lymphocyte growth transformation. EBV strains with deletion mutations in the LMP-1 gene were found to be incapable of immortalizing B lymphocytes.52 LMP-1 has been shown to be responsible for many of the phenotypic changes observed in EBV-infected B-lymphocytes including upregulation of expression of B-cell activation markers (CD21 and CD23), the cell membrane adhesion molecule ICAM-1, and the bcl-2 protooncogene.53 Increased expression of adhesion molecules facilitates cell clumping which may enhance B-cell growth and proliferation via paracrine growth factors. T cells also adhere to B cells expressing these adhesion molecules, and such T-cell adherence may represent the initial step in the host immune response to EBV infection that ultimately leads to elimination of EBV infected lymphoblasts in vivo.54
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


