The Ciliary Body
Ann Barker-Griffith
Barbara W. Streeten
Michael Fikhman
Justin Gutman
The functions of the ciliary body are among the most diverse of any ocular tissue, since it secretes the aqueous, nourishes the lens, and provides the muscle power for accommodation, contributing at the same time to control of aqueous outflow. It also forms part of the vitreous base, serving as an attachment for and probably synthesizing the unique zonular fibers, and has also been suggested as a source of vitreous hyaluronic acid.1,2,3
SURFACE LANDMARKS AND RELATIONSHIPS
The shape of the ciliary body is a long triangle in cross section, its anteroposterior length varying considerably with the length of the eye (Fig. 13-1). It is always longer on the temporal side and longest inferotemporally, the most expanded quadrant of the eye. Temporally it is 5.6 to 6.3 mm long and nasally 4.6 to 5.2 mm.4 Of importance surgically is the distance from the external corneoscleral limbus to the end of the ciliary body at the ora serrata. This may be roughly estimated as 7 mm temporally and 6 mm nasally, since in 200 autopsy eyes the slightly shorter distance from the Schwalbe line internally to the end of the ciliary body at the ora serrata averaged 6.53 mm temporally, 5.73 mm nasally, 6.14 mm superiorly, and 6.20 mm inferiorly.5 However, variation in ocular length can be considerable, such as in buphthalmos where the temporal ciliary body may be up to 10 mm long. Another variable is increase in thickness of the ciliary body with increasing myopia and axial length, as measured by ultrasound microscopy.5 The anterior face of the ciliary body forms a 45° to 90° angle with the scleral surface (Fig. 13-1), depending on age and accommodative state.6 It is directly exposed to the aqueous, forming part of the posterior boundary of the anterior chamber and filtration angle. The iris root arises from the midportion of this anterior face. At its anterior end, the ciliary body is attached to the scleral spur region, its firmest point of fixation to the globe, and its connective tissue blends into that of the trabecular meshwork. Along the rest of its outer surface, the ciliary body has loose attachments to the sclera, separating readily during processing for microscopy. This ciliary-scleral junctional space is called the supraciliaris or supraciliary lamina.
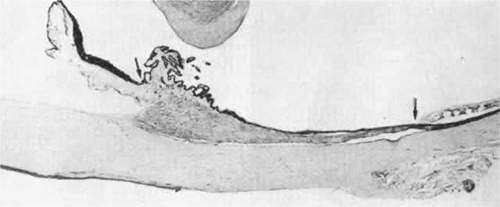 Figure 13.1. Cross section of the ciliary body, extending between the arrows, in a 64-year-old human eye with the retina at the ora serrata (right arrow) and the anterior face of the ciliary body (asterisk). The edge of the lens lies above the ciliary processes (pars plicata) and anterior ciliary body. (Hematoxylin and eosin, X 20.) |
When the intact globe is transilluminated (Fig. 13-2), light is visible coming through the pupil and the posterior two thirds of the globe with the posterior boundary between ciliary body and retina at the ora serrata (OS) is usually well seen, as the pigment epithelium on both sides of the ora serrata is heavily pigmented, forming a 1.5- to 2-mm dark band. The anterior 1 to 1.5 mm of this band belongs to the ciliary body, with a clear space indicating the pale pars plana (PP) separating it from the pigmented pars plicata and iris. Visibility of the pars plana by this method can be a useful landmark indicating an entry site when needed, through the pars plana rather than the retina, for intraocular surgery or photocoagulation procedures.
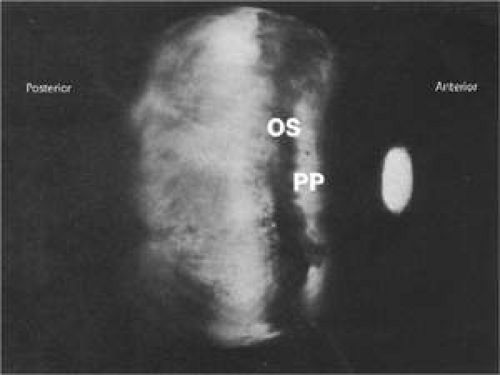 Figure 13.2. Transscleral transillumination through an intact globe showing the darkly pigmented ora serrata (OS) region contrasting with the paler pars plana (PP). |
When a globe is opened through its equator (Fig. 13-3), the internal anterior view shows the posterior white lens centrally surrounded by a wreath of white ciliary processes and an outer dark rim of ciliary epithelium (Fig. 13-3A). To the early anatomists this striking appearance resembled white eyelashes (cilia), a fanciful analogy perpetuated in the name of this muscular structure.1 The ciliary processes are pigmented early in life but gradually lose pigment with aging. These elevated processes are known as the pars plicata, and the flat area from their endings out to the ora serrata (OS) is known as the pars plana. It is clear in Figure 13-3A that the dark pars plana is broader on the temporal (T) (right) side than it is on the nasal (N) (left) side. Transillumination of the tissue in Figure13-3B shows more clearly the same difference in widths of the nasal (N) and temporal (T) pars plana, while the thick ciliary processes appear only as a dark ring around the lens. Also visible is that the whole pars plana is divided into segmental radial units, separated by thin hyperpigmented ridges. These radial units have scalloped outer bases on the nasal (N) side and flattened bases on the larger temporal (T) side.
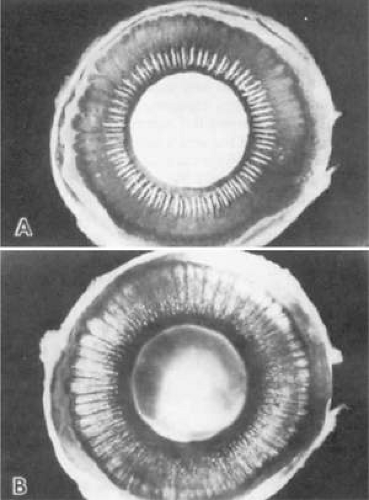 Figure 13.3. Posterior view of inner surface of the ciliary body, with the lens in place surrounded by white ciliary process. A. The nasal side (N) is shorter than the temporal side (T) and has a scalloped cilioretinal junction. B. Transillumination of above eye demonstrates division of the whole ciliary body into segmental units of pale pars plana (white arrow), outlined by radially arranged hyperpigmented ridges. |
Seven anterior ciliary arteries enter the ciliary body from the outer rectus muscles through scleral canals just behind the pars plicata. These entry sites are the only other firm attachments of the ciliary body to the outer globe. In 12% of eyes, a loop of long posterior ciliary nerve enters the sclera from the ciliary body in this region, called a nerve loop of Axenfeld.7 Sometimes the same canal holds an anterior ciliary artery as well. These loops seem to have no specific purpose and return to continue their anterior course in the ciliary body (Fig. 13-4). They may be visible subconjunctivally 3 to 4 mm from the limbus, especially in the inferior quadrants. When associated with melanin pigment, the loop may appear as a pigmented spot under the conjunctiva, requiring differentiation from a rare extraocular extension of a malignant melanoma as seen in Figure 13-4. Sensitivity of a nerve loop to touch and absence of a tumor shadow on transillumination can aid in this diagnosis.
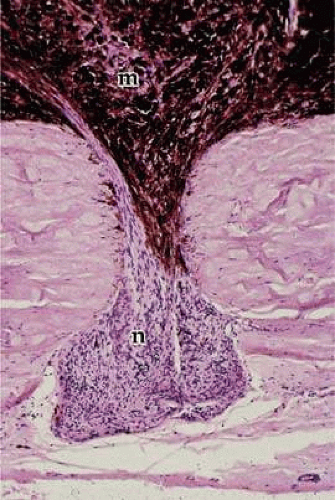 Figure 13.4. Nerve loop of Axenfeld (n) extends into a scleral canal along with an anterior ciliary artery (not visible here). A melanoma (m) is invading the same scleral canal, an uncommon occurrence. (Hematoxylin and eosin, × 120.) |
The posterior limit of the ciliary body is sharply demarcated on its inner surface by its abrupt junction with the retina at the ora serrata. No such demarcation occurs on its outer surface, where the supraciliaris blends directly into the suprachoroidalis. The inner surface of the ciliary body has a close relation to the zonular fibers, except for the anterior rounded ends of the ciliary processes. On its posterior half, the inner ciliary surface gives attachment to the anterior two thirds of the vitreous base, and its anterior half forms the lateral boundary of the posterior chamber. The lens lies 1 to 1.5 mm centrad to the ciliary processes, depending on age and lens diameter, as the enlarging lens narrows the perilenticular space.
The ciliary body is easily freed from the sclera by a spatula passed through its attachment to the scleral spur, allowing inspection of both supraciliary surfaces. The scleral surface is smooth, slightly pigmented, and marked only by canals for ciliary arteries and nerves. The ciliary surface has loose sheets of pigmented stroma and muscle over its posterior two thirds. On these lie the thin, long ciliary arteries and the flat, ribbonlike long ciliary nerves, both of which may bifurcate at the posterior border of the ciliary body (Fig. 13-5). The anterior third of this surface is marked by the slightly convex white mass containing the main bulk of the ciliary muscle (Fig. 13-5A). On transillumination the muscle appears dark (Fig. 13-5B) as does the ora serrata, with pale pars plana between, similar to the view in Figure 13-2.
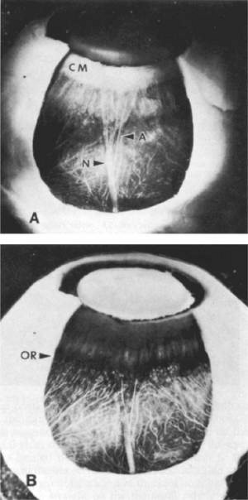 Figure 13.5. Outer surface of ciliary body seen after removal of a scleral window, cornea, and iris. A. Most of the ciliary muscle (CM) lies in the anterior half as a white band. A flat ribbonlike long posterior ciliary nerve (N) and thin artery (A) bifurcate shortly before the ora serrata. B. Transillumination indicates the actual site of the ora serrata (OR) and relationships to the choroidal vessels. |
THE ANTERIOR CILIARY BODY
In the fetus, development of the ciliary body can first be recognized in the fourth month by elevation of thin radial folds from the smooth surface of the anterior optic cup around the lens. These folds continue to enlarge, thicken, and push into the posterior chamber. The anterior ends protrude forward like rounded beaks, increasing the surface area that will secrete aqueous (Figs. 13-6A and 13-6B). These 70 to 80 folds become the ciliary processes and are collectively known as the corona ciliaris or pars plicata portion of the ciliary body. The pars plicata is usually 2 mm in anteroposterior length in the adult. Each process is about 0.5 mm in width, may reach 1 mm in height, and shows a fine rugation in the coronal plane. An exact count of the processes cannot always be made because of frequent joining of their anterior ends and development of minor processes or plicae in the valleys between them. These sausagelike minor plicae are prominent in the posterior third of the pars plicata. Giant processes are often found in the nasal horizontal meridian.
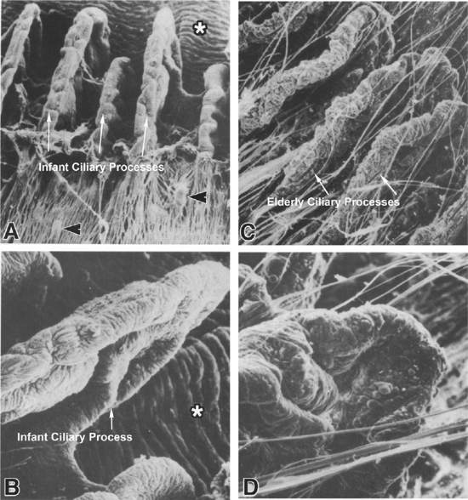 Figure 13.6. Scanning electron microscopy of the ciliary processes of an infant and adult. A and B. Age 3 months. The zonules have been rolled back to reveal the simple infantile processes. Several minor plicae (arrowheads) are visible. The peripheral iris folds are seen above (asterisk). (A, × 60; B, × 115) C and D. Age 85 years. The processes are larger, more convoluted, and warty. (C, × 45; D, × 110.) |
The ciliary processes continue to enlarge and become more convoluted in adult life, gradually losing superficial pigment so that their crests are whitened (Fig. 13-3A). A marked knobby or warty change develops on their surfaces and in the valleys between them in old age (Fig. 13-6C and 13-6D). In the infant, the fine zonular fibers are draped over the rudimentary ciliary process region like a smooth, silvery membrane. As the processes elevate and become more complex, the zonules remain attached in the valleys between them and along their sides, dividing them into separate aggregates of fibers (Fig. 13-6C), with few passing over the heads of the processes, allowing free entry of aqueous into the posterior chamber. The anterior beaklike ends of the processes are always free of zonular fibers.
THE CILIARY SULCUS
A surface anatomic region of considerable interest anteriorly is the ciliary sulcus, which is the valley created under the iris at its origin from the ciliary body. This sulcus is bounded posteriorly by the ciliary processes and more centrally by the anterior zonular fibers and the lens surface (Fig. 13-7A), making it a natural site for supporting the haptics of a posterior chamber intraocular lens (IOL). Although the ideal site for a posterior chamber IOL is within the lens capsular bag, such placement is not possible when the lens capsule is absent or where the capsule-zonular diaphragm lacks sufficient strength, such as in pseudoexfoliation when some of the zonular fibers have ruptured. To choose an appropriate size of IOL for support and centration in this space, it is important to consider the diameter of the sulcus. In 41 postmortem eyes this diameter correlated significantly with axial length determined echographically, with a mean sulcus diameter of 11.2 mm (SD = 0.3 mm).8 A bimodal distribution of diameters was noted with a clustering of smaller diameters in shorter eyes. It was suggested that this results from incomplete posterior migration of the pars plicata from the peripheral iris during embryonic growth. In another postmortem study,9 the mean sulcus diameter was slightly smaller than the mean corneal diameter. In contrast, a study of 64 human autopsy eyes by Blum et al10 showed that there was no age correlation in the horizontal, vertical, longitudinal ocular measurements, and corneal diameters. The vertical ciliary sulcus diameter decreased from 12.02 ± 0.12 mm in the ≤40-year-old or younger group to 10.71 ± 0.91 mm in the ≥80-year-old or older group. The horizontal ciliary sulcus diameter decreased from 11.36 ± 0.24 mm in ≤ the 40-year-old or younger group to 10.33 ± 0.76 mm in the ≥80-year-old or older group.10 Pop et al11 found that sulcus size could not be predicted in 43 eyes using traditional limbal measurements owing to variation in axial lengths, causing a 0.6 mm error for myopia and a 0.3 mm error for hyperopia. Since the limbus size does not correlate with age, it alone is not adequate enough to predict the diameter of the ciliary sulcus. Instead, Pop et al11 suggest that a general formula incorporating the mean keratometry and sphere can help predict the size of the ciliary sulcus:
Sulcus size = 18.9 + (-0.023 x sphere) + (-0.15 x mean keratometry)
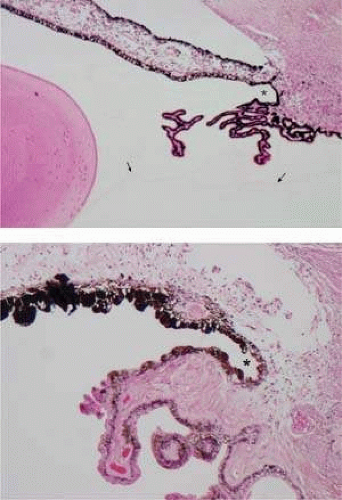 Figure 13.7. The ciliary sulcus. A. Cross section through a ciliary sulcus (asterisk) of a child. Arrows indicate anterior hyaloid of vitreous. Lens is on left. B. Cross section through a ciliary sulcus of a 51-year-old patient. Irregular iris pigment (above) and ciliary nonpigment epithelial cells line the sulcus (asterisk). (Hematoxylin and eosin, A, × 43; B, × 165.) |
Smith et al12 also noted that the mean distance from the corneal limbus to the ciliary sulcus measured externally was 0.9 mm, which would be of interest for transscleral suture fixation of haptics.
The bands and folds that sometimes extend between ciliary processes and posterior iris can misdirect haptics into the easily penetrated iris or ciliary tissues during IOL insertion.13 It is critical to determine the correct lens size to place into the ciliary sulcus because failure to do so could cause secondary glaucoma from the release of iris pigmentation and structural damage to the ciliary body. This has led to recommendations for the use of IOL specifically designed for the placement into the ciliary sulcus.13,14,15
THE PARS PLANA AND ORA SERRATA
The posterior portion of the ciliary body, known as the pars plana or orbiculus ciliaris, is about 4 mm long on the temporal side and 3.5 mm on the nasal side, depending on the length of the eye. It is limited posteriorly by its junction with the retina at the ora serrata. There is considerable variation in configuration of this junction, but generally it is straight on the temporal side and distinctly scalloped on the nasal side (Fig. 13-3). In the infant there is no scalloping, and in some adults scalloping may extend all around the ora serrata.
Transillumination of the ciliary body demonstrates a segmentation into radial ciliary units (Figs. 13-3B and 13-8). A ciliary process lies in the center of each unit, whose sides are demarcated by pigmented ridges called striae ciliaris. These striae extend almost to the pars plicata and often have minor processes or plicae at their anterior ends. The nasal ora serrata demonstrates ciliary unit formation more clearly than does the temporal side (Fig. 13-8). The oral concavity opposite the central ciliary process is known as a ciliary bay, limited on each side by extension upward of retinal “teeth,” also called dentate processes. These average 16 per eye5 and extend up over the pigmented ridges. Sometimes two bays may join when there are large accessory ciliary processes or plicae. Giant dentate retinal processes occur aberrantly in 16% of eyes,16 directly onto the ciliary processes rather than between them, and may merge with the process. Such large “meridional complexes” usually include an elevated meridional fold of tissue composed of anomalous retina and ciliary epithelium (Fig. 13-9).4,16 These anomalies are frequently associated with aberrant zonular bundles, with small holes or shallow retinal excavations behind the complexes.
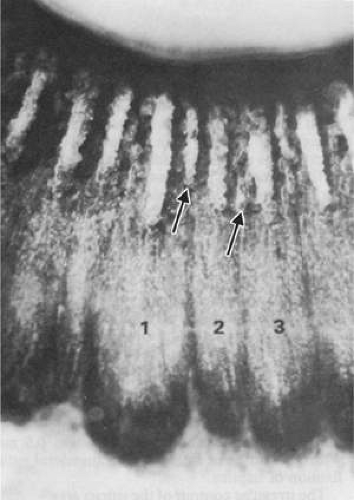 Figure 13.8. The ciliary process unit. Three ciliary units on the nasal side of the ciliary body. Pigmented ciliary ridges demarcate the edges of the unit, with a ciliary process lying centrally. Minor processes and plicae tend to occur at the ends of the ridges (arrows). (Unstained × 15.) |
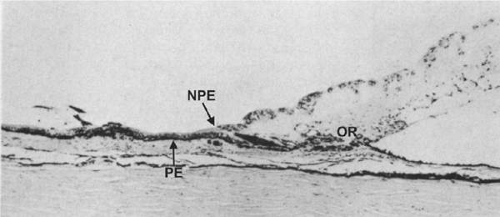 Figure 13.9. Meridional fold showing anomalous development of the retina and ciliary epithelium at the ora serrata (OR). (Hematoxylin and eosin, × 80.) |
The two layers of ciliary epithelium are firmly adherent to each other and to their underlying stroma at the ora serrata, where the nonpigmented layer is suddenly converted into retina. The more loosely attached retina with any retinal teeth can be stripped from this firm junction by vitreous traction, creating the sharply demarcated retinal edge seen in traumatic disinsertions of the peripheral retina.
The dense attachment of vitreous known as the vitreous base extends from the peripheral retina up onto the posterior 2 mm of the pars plana, with less dense adhesion up to its separation as the anterior hyaloid membrane. A grayish line frequently appears just behind the anterior limit of vitreous attachment (Fig. 13-10A), visible clinically with a three-mirror lens. This gray line is the edge of one of the vitreous lamellae that develop in adults, separating the vitreous cavity into several anteroposterior compartments. Eisner17 first described these compartments, referring to them as tracts (Fig. 13-10B). The lamella inserting on the mid–pars plana was named the median tract. One or more over the ciliary processes constituted the coronary tract, and that at the periphery of the lens the anterior hyaloid tract. Circumferentially arranged zonular and vitreous girdle fibers can be seen on and within the anterior hyaloid membrane, appearing to support the vitreous attachment to the ciliary body and lens during ocular movement and accommodation.18
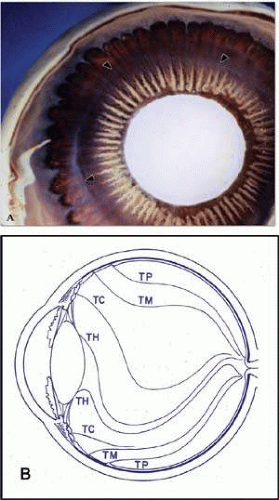 Figure 13.10. Vitreous attachments to ciliary body. A. Circular reflections (arrowheads) from the vitreous lamellae or tracts present in the otherwise invisible vitreous as they curve upward over the ciliary processes. B. Diagram of tracts dividing the vitreous of the adult into several anteroposterior compartments. TP, preretinal tract; TM, median tract; TC, coronary tract; TH, hyaloid tract. (B, Eisner G: Clinical examination of the vitreous. Trans Ophthalmol Soc UK 95:360, 1975.) |
MICROSCOPIC ANATOMY OF THE CILIARY BODY
NONPIGMENTED CILIARY EPITHELIUM
The ciliary epithelium consists of two layers: an inner nonpigmented (NPE) and an outer pigmented layer (PE). These two layers derive from infolding of the single cell layer of the optic vesicle against itself, to form the optic cup. The potential space left between the two ciliary layers rarely opens, owing to the frequency of junctional complexes uniting the cells. A peculiar result of the infolding affects nomenclature in this region, since the apices of the epithelial cells now face each other across a potential space. The bases of the cells face outward, toward the ciliary body stroma for the PE, and toward the posterior chamber for the NPE (Fig. 13-11). Basement membrane covers the bases of both cell layers as is characteristic of epithelial cells.
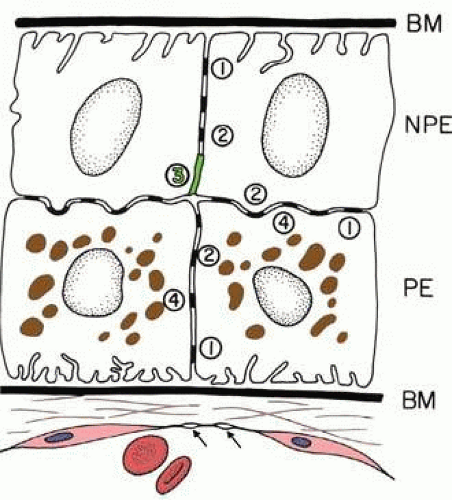 Figure 13.11. Diagram showing the relationship of the two layers of ciliary epithelium. Basement membrane (BM) covers their bases, and their apices face each other across a potential space. The cells are united by several types of junctional complexes: 1, desmosomes; 2, gap junctions; and 4, puncta adherentes. Only the apices of the nonpigmented epithelial cells (NPE) are joined by tight junctions (3, shown in green), composed of a zonula occludens usually combined with a zonular adherens complex. PE, pigmented epithelium. Arrows show capillary fenestrations. |
The NPE of the ciliary body stretches in a continuous layer from the root of the iris to the ora serrata. As the transition from pigmented iris epithelium occurs, melanin granules in the inner layer suddenly decrease in number, and the cells become slightly smaller (Fig. 13-12). In the anterior pars plicata the NPE cells are cuboidal, 12 to 15 μm in width, with central nuclei and in the posterior pars plicata there is some pigment epithelial thickening. The knobbiness that develops during aging is due to small nodular surface proliferations of NPE cells, especially on the minor plicae (Fig. 13-13A). In the young eye the cells of the pars plana are also cuboidal, but with growth become thinner and more columnar, sometimes reaching up to 30 μm in height and 5 to 10 μm in width. In the posterior half of the pars plana, some of the NPE cells tilt forward as though responding to anterior zonular traction, while others may be inclined posteriorly, suggesting complex vectors of force in this region (Fig. 13-13B). The nuclei are vertically oval and lie near the apex of the cells. The epithelium here becomes very irregular with aging, showing hyperplastic toothlike cell processes intertwining and extending up into the vitreous and among the zonular fibers. At the ora serrata the ciliary NPE may join the retina abruptly, highlighting the difference in thickness of these two layers (Fig. 13-14).
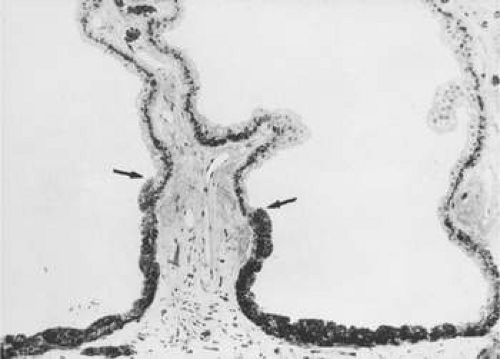 Figure 13.12. Frontal view of a ciliary process at its junction with iris, showing conversion to thicker, double-layered iris pigment epithelium (arrows). (Toluidine blue, × 200.) |
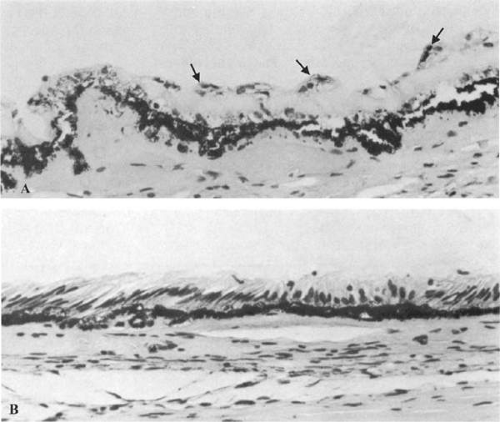 Figure 13.13. Ciliary nonpigmented epithelium. A. Nodular proliferation of the nonpigmented epithelium appears as small cellular caps over the surface at age 70 years. Z, zonule. (Hematoxylin and eosin, × 400.) B. Distorted epithelium of pars plana shows evidence of traction in both anterior and posterior directions (adult). (Hematoxylin and eosin, × 400.) |
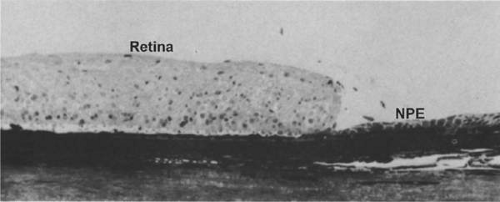 Figure 13.14. Ora serrata in young adult, showing abrupt junction of ciliary nonpigmented epithelium and sensory retina. (Toluidine blue. × 200.) |
Whereas the ultrastructure of the PE is similar in most areas of the ciliary body, that of the NPE shows noticeable regional differences that appear to be of functional importance. These differences have been studied more extensively in the monkey than in the human but are similar in both species.19,20,21,22,23,24,25,26,27,28,29,30 The pars plicata, and particularly its anterior portion, appears to be the predominant site of aqueous formation and has many special characteristics of this secretory function. The NPE here has marked cytoplasmic infolding at its base and redundant interdigitations at its basolateral margins, greatly increasing the area of cell surface facing the posterior chamber (Fig. 13-15, inset A). These cell membranes and to a lesser degree those of the pigmented epithelium, contain the enzyme complex Na+/K+-ATPase,33 evidence of an energy-dependent active transport system.31 The presence of the enzyme carbonic anhydrase in the NPE cells of the pars plicata of all species studied is further evidence of fluid-pumping activity.32 In the moderately electron-lucent cytoplasm of the anterior NPE particularly, there are many mitochondria near the base of the cell and rough endoplasmic reticulum (RER) in single cisternae or parallel stacks near the nucleus, where Golgi complexes are also common (Fig. 13-16). The mitochondria are important to provide energy for transport, and the RER for processing of new protein. Clusters of free ribosomes and occasional cilia are found in all areas. With aging, unusual whorled formations of RER are described near the cell base in the pars plicata, along with lipid droplets and lysosomal residual bodies.20,25
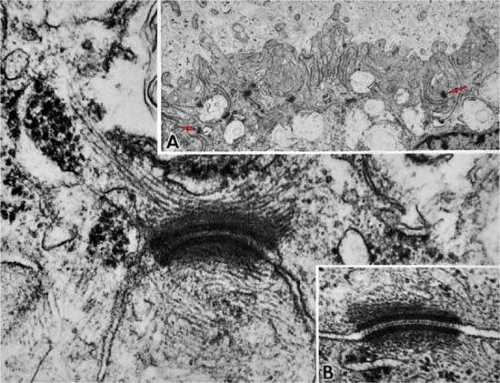 Figure 13.15. Anterior pars plicata. Inset A. The basement membrane does not enter the extensive surface infoldings and lateral interdigitations of the nonpigmented epithelium. Note large numbers of desmosomal intercellular junctions (arrows). Main figure. A desmosome cut slightly obliquely showing intermediate filaments of the cell’s cytoskeleton inserting into the dense plaque material. Inset B. Central dense core and plasmalemmal unit membranes of desmosome cut perpendicularly. (Main figure and inset B, × 104,000; inset A, × 10,800.) |
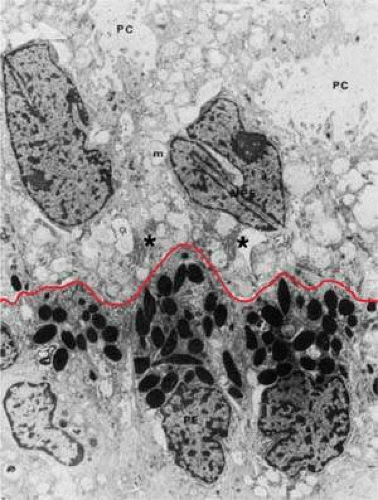 Figure 13.16. Ciliary epithelium in the anterior pars plicata of a 19-year-old. Posterior chamber surface (PC) of the nonpigmented epithelium (NPE) has a sawtooth pattern (red line) seen irregularly throughout this layer with aging. Mitochondria (m) are large, plentiful, and show artifactual hydropic change. Rough endoplasmic reticulum is present below the nuclei (asterisk). Note that the apices of the pigmented epithelial cells (PE) are conical, with prolongations of NPE between them. (× 8,320.) |
In the child’s eye, the basement membrane is a simple thin layer over the base of the NPE cells, not extending into the surface or lateral infoldings (Fig. 13-17, inset A). As is typical for thin basement membranes, it is a 30-nm granular layer separated from the plasmalemma by a 50-nm lucent zone. Beginning in the first decade, the basement membrane undergoes a remarkable thickening of the multilaminar reticulated type. This change has been identified by the age of 3 years,20 beginning in the valleys of the posterior half of the pars plicata. From this region, thickening by multiple intertwining thin layers of reticular basement membrane extends posteriorly and up onto the lateral walls of the ciliary processes, involving almost all of the ciliary epithelium in those older than 50 years of age. The thickened basement membrane may reach 2 μm with an increasing quantity of membrane-bound vesicles and granular material, apparently debris of cellular metabolism by age 20 years, whereas at age 7 years the basement membrane was very thin with minimal multilamina change (inset A). By scanning electron microscopy the basement membrane surface had a fibrogranular mossy appearance already present (inset B). Zonular bundles are seen in close apposition to the basement membrane and superficially within it in the ciliary valleys, to which they have a very firm attachment.l8
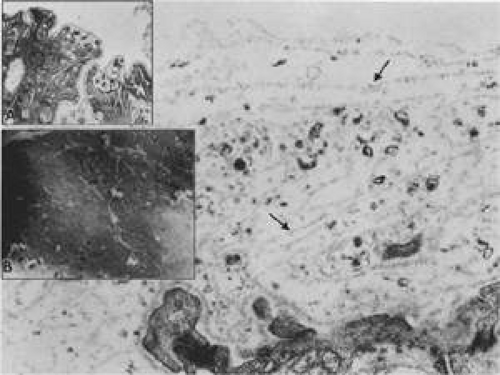 Figure 13.17. Changes in the ciliary nonpigmented epithelial basement membrane (arrows) that occur with aging. Main figure shows multilaminar pattern of basement membrane proliferation filled with vesicles and granular debris at age 20 years. (× 54,600.) Inset A. Thin basement membrane with patchy multilaminar change at age 7. (× 15,000.) Inset B. Scanning electron micrograph showing fibrogranular mossy appearance of basement membrane surface over sides of anterior ciliary processes at age 20 years. (× 10,200.) |
In the posterior pars plana, the tall NPE cells have numerous intermediate filaments and granules that give the cells a more electron-dense appearance than anteriorly (Fig. 13-18A) and are profusely supplied with tubules of smooth-surfaced endoplasmic reticulum (Fig. 13-18B). The intermediate filaments in the NPE of the pars plicata over the ciliary crests and in the pars plana are strongly immunoreactive for vimentin, with fainter staining for cytokeratin 18, just the reverse of the staining pattern in the ciliary PE.29 The vimentin-positive intermediate fibers attach the cytoskeleton to adherens junctions and often indicate cells subject to tractional or cell-shape stresses. Fine actin filaments are present in the cytoskeleton of both epithelial cell types, without regional differences. Extensive cystic dilatations of the intercellular spaces between the NPE cells commonly occur in the posterior pars plana of adult eyes (Fig. 13-18). Fine and Zimmerman2 showed that these spaces contain hyaluronidase-sensitive acid mucopolysaccharide and suggested it might be hyaluronic acid intended for the vitreous. An increased number of Golgi complexes in this region also suggests production of some glycoprotein.25 Immunostaining for the membrane-bound enzyme hyaluronan synthase is positive on the membranes of the primate posterior pars plana NPE cells and hyalocytes, consistent with local hyaluronan secretion, as reported by Lutjen-Drecoll.3 Interestingly, staining is equally intense over the crests of the ciliary processes.
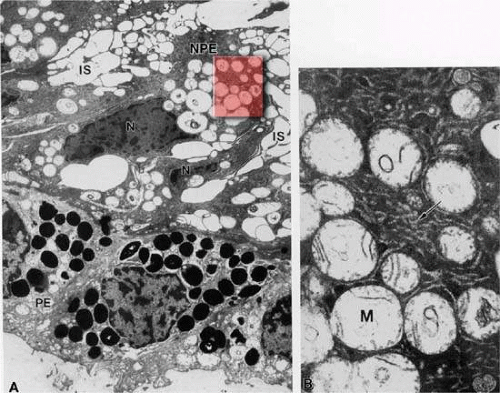 Figure 13.18. Posterior pars plana. A. Nonpigmented epithelial (NPE) cells appear tilted and compressed with dense cytoplasm. The extensive cystic dilatation of the intercellular spaces (IS) around these cells is characteristic of this region. Only the nucleus (N) and the many hydropic mitochondria are visible in the cytoplasm. The basement membrane under the pigment epithelium (PE) is multilaminar and thick. (× 5,700.) B. Higher magnification of highlighted region in A shows that the cytoplasm is full of branching and curved profiles of smooth endoplasmic reticulum (arrow), which is also typical of this region. M, mitochondrion. (× 22,000.) |
There are many intercellular junctions between the ciliary epithelial cells, each giving important data about the specific functions of these cells. The most common is the desmosome, seen well in Figure 13-15 as focal dark areas in Figure 13-15 (inset A) binding the plasma membrane of adjacent cells together, leaving a narrow clear zone between the edges of the cells with dense plaquelike material on each side. The lateral sides of two cells are usually joined by several interrupted areas to fulfill a structural need. Secretory cells have a primary need for control of fluid flow within them and require tight junctions at the apical ends to control flow of material, consisting primarily of a zonular occludens with a zonular adherens above it. These tight junctions represent the primary blood-aqueous barrier in the ciliary body. When large tracer molecules such as horseradish peroxidase are injected intravenously into primates,23,24 the tracer has an easy passage through the fenestrated capillaries of the ciliary processes but cannot pass beyond the apices of the NPE cells, as can also be shown by much smaller tracers (Figs. 13-19 and 13-20). Since the tight junction is a site where the plasmalemmal membranes of the adjoining cell surfaces are adherent, freeze fracture technique can split these apart demonstrating the very fine dots on the P face of one of the cells, which help it adhere to the outer plasmalemmal leaflet. Looking at the outer leaflet shows a regular quilted pattern binding the two faces together. The number of these quilted strands determines how tight the junction is intended to be, according to the need for virtually a complete closure (Fig. 13-21). The separate zonular occludens accompanying the adjacent zonular adherens is usually arranged in the band form around the upper end of the tight junction. Another important junction is the gap junction, which occurs in “extraordinary numbers” in the ciliary epithelium and may be found even among the strands of zonula occludens24 (Fig. 13-22). At gap junctions, the surface membranes of the two cells run a very straight course, separated by only a 2- to 3-nm cleft, and are best seen as a dark prominent line attaching to surfaces of the nonpigment and pigment epithelium. This strong junction appears like rounded areas of spot welding when seen flat with very fine nodules over the surface and may have some function in transfer of small molecules across the junctional space, occurring in very large numbers in the ciliary epithelium. These numbers indicate that the cells are closely coupled for electrical and metabolic cooperative work across the sites of low ionic resistance.
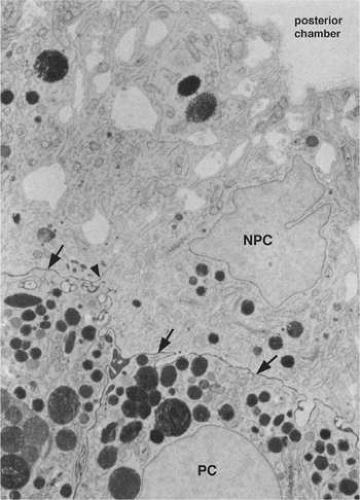 Figure 13.19. Evidence of tight junctional complexes in the anterior ciliary epithelium of Macaca mulatta. The pigmented epithelial cells (PC) are outlined by a thin black line (arrows) that is the reaction product of horseradish peroxidase, a tracer substance injected intravenously in vivo. The tracer has entered the intercellular spaces of the nonpigmented epithelium (NPC) but is held up by occluding junctions (arrowheads), preventing further progress into the posterior chamber. (× 8,450; courtesy of Dr. Guiseppina Raviola.) |
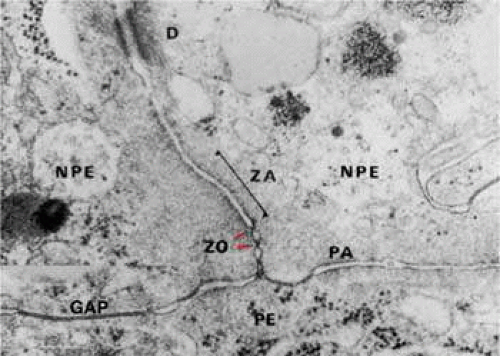 Figure 13.20. Apical junction of two nonpigmented epithelial cells (NPE) showing focal zonula occludens (ZO) junctions (red arrowheads), an adjacent zonula adherens (ZA), and desmosome (D), puncta adherentes (PA), and gap (G) junctions. PE, pigmented epithelium. (× 58,700.)
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
 Get Clinical Tree app for offline access
Get Clinical Tree app for offline access

|