Olfaction
Eric H. Holbrook
Donald A. Leopold
Anyone who has experienced complete nasal obstruction from an infection or severe allergic rhinitis has suffered the loss of a sense that is often taken for granted. In other mammals, the sense of smell is absolutely crucial for survival, reproduction, and rearing of young. Although humans can survive without olfaction, the negative impact on quality of life has been well documented with some approaching clinical depression (1,2,3). As other animals depend on the sense of smell as an alarm system for danger, we also depend on the sense of olfaction to warn us of smoke, natural gas leaks, and spoiled food. On a more frequent occasion, people with smell loss struggle with the blandness of meals and have difficulty with the appropriate use of bodily hygiene products or perfumes. In the food industry, olfaction is crucial to chefs and wine tasters and its loss can be devastating.
The overall prevalence of olfactory dysfunction includes 19% of the population over the age of 20 and 25% of the population over the age of 53 (4,5). If smell loss from aging alone is considered, one out of eight people between the ages of 53 and 91 will acquire an olfactory impairment over a 5-year period (6). Many patients will report a loss in taste when in reality they are experiencing a decrease in flavor due to a lack of sense of smell. Regardless, the effect is the same and especially in the elderly population, the impact on diet and nutrition can be significant (7,8).
This chapter describes the main causes of olfactory disorders and provides a logical guideline to the evaluation and treatment (when available) of patients with smell dysfunction. The following commonly used definitions describe olfactory disorders. The complete absence of a sense of smell is termed anosmia, while hyposmia is a decreased sense of smell. Dysosmias are alterations (decreases or distortions) in smell. A smell perceived in the absence of a true odor in the environment is termed phantosmia, while an altered perception of an odor in the environment different from what is usually experienced for that odor is termed parosmia.
ANATOMY
The nasal cavity is the conduit for odors to reach the specialized neuroepithelium that converts binding of odorant molecules by receptors into electrical signals that extend to the brain. Many studies have looked at airflow properties of the nasal cavity and ability to smell. Although severe nasal obstruction from septal deviation can reduce odorant access to the olfactory nerves, it is rarely the sole cause of olfactory loss. Retronasal presentation of odors during eating has been given increasing importance for adding separate qualities to the overall appreciation of flavor of food (9,10).
Ultimately, odorants need to interact with olfactory receptors that reside in a patch of olfactory epithelium located in the superior/posterior-most aspect of the nasal cavity, the olfactory cleft. This area is variable in size and variable in the extensiveness of small patches of respiratory epithelial replacement. The overall area averages 1 to 2 cm2 in adults, but covers a much larger region in infants (11). The decrease in olfactory epithelium over time and the presence of respiratory patches within the olfactory epithelium is speculated to be caused by cumulative environmental insult and subsequently a source for olfactory loss with aging. The olfactory epithelium covers a region of the septum below the cribriform plate extending back to the face of the sphenoid (12). A similar sized region apposed to the septal olfactory mucosa is present laterally within the olfactory cleft on the middle and superior turbinates. The narrowness of this space may protect the delicate olfactory neurons from environmental damage, but may increase susceptibility to obstruction from inflammatory processes with resulting smell loss.
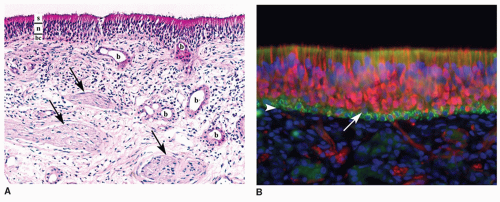 Figure 24.1 Human olfactory mucosa. A: Hematoxylin and eosin stain of human olfactory mucosa (200×). The epithelium can be divided into a basal cell layer (bc), neuronal layer (n), and supporting cell layer (s) where the cell bodies of each of these cell types typically reside. The underlying lamina propria contains Bowman glands (b) and fila olfactoria (arrows) with multiple olfactory receptor axons. B: Immunohistochemical staining of human olfactory epithelium (400×). Olfactory neurons with their dendrites and axons are labeled red with an antibody directed against PGP9.5. The fine dendrites are seen extending between supporting cells and ending in knobs at the apical surface, while the axons exit through the basal lamina (white arrowhead) to converge and join the fila olfactoria. Horizontal basal cells (HBCs) are labeled green with antibodies against keratin 5 and reside on top of the basal lamina. Unstained cells between the HBCs and neurons are likely globose basal cells (arrow). Supporting cell nuclei are seen as the row of blue (DAPI stained) nuclei above the neuronal layer. |
The olfactory mucosa is composed of a pseudo-stratified neuroepithelium (Fig. 24.1). Supporting cells populate the apical portion of the epithelium and send foot processes to the basal lamina. The dendritic knobs of the olfactory neurons with their multiple immotile cilia extend above the apical surface of the supporting cells, but the cell bodies of the neurons reside in the midsection of the epithelium with the more mature neurons taking a more apical position. The axons extending from these bipolar cells extend through the basal lamina and converge with other olfactory axons to form bundles (fila olfactoria) that transverse the cribriform plate and innervate the brain within the olfactory bulbs at synaptic collections termed glomeruli. These axons are unique in several aspects: they are true receptor neurons with direct synapses to the brain, they are the only cranial nerve (CN I) with direct exposure to the environment, and they are continually replaced throughout our lifetime. Signal projections from the olfactory bulb travel through the lateral olfactory tracts and synapse on several higher locations including the anterior olfactory nucleus, amygdala, piriform cortex, and entorhinal cortex. Major projections to structures associated with the limbic system and hippocampus explain the often-experienced relationship of specific odors with a strong emotional response or triggering of a vivid memory associated with the odor.
The cells responsible for the replacement of these neurons as well as replacing other cells comprising the epithelium are the horizontal and globose basal cells positioned just above the basal lamina. Unique to the olfactory mucosa are Bowman glands within the lamina propria that send ducts through the epithelium to the apical surface providing mucus that is likely important in the presentation of odorants to the olfactory receptors. Although there is variation in the thickness of the epithelium and less ordered positioning of the olfactory neurons within the epithelium compared to that found in rodents, the architecture and cellular components are remarkably similar in humans (12).
PHYSIOLOGY OF OLFACTION
The detection and identification of a specific odor from the multitude of complex odors present in the environment is only partially understood. The initial site of interaction between an odorant molecule and the olfactory neuron, the olfactory receptor, has now been identified (13). These receptors collectively form a family of similar G proteins (Golf) that when activated stimulate adenylate cyclase to synthesize cyclic adenosine monophosphate (cAMP). cAMP acts as a second messenger to open a cyclic nucleotide-gated ion channel allowing for influx of cations and depolarization of the neuron. There are approximately 1,000 different genes within the olfactory receptor gene family in rodents; many however are pseudogenes. The human olfactory gene family with 339 intact receptor genes and 297 pseudogenes has fewer receptor genes than mice; however, this gene family still comprises one of the largest
gene families in the human genome (14). In rodents, it has been shown that each olfactory neuron expresses only one member of the olfactory receptor genes. Neurons expressing like receptors are restricted to a regional zone oriented from a dorsal-medial to ventral-lateral orientation; however, within these regions, the expression pattern appears random among other receptors expressed in the similar area. These neurons expressing like receptors extend axons that remarkably converge together within the lamina propria to eventually converge exclusively to, on average, two glomeruli within each olfactory bulb (15). In humans, this receptor-to-glomeruli ratio is much broader and calculated to be on the order of 1:16 (16). This receptor-specific patterning is the basis of an odorant map whereby an odorant stimulates a subset of olfactory neurons that in turn activate a specific set of glomeruli in the olfactory bulb (17,18). Therefore, an odor is coded by the multiple receptor neurons it stimulates and in turn the resulting unique pattern of glomerular activation is transmitted in an unclear fashion to higher brain centers and perceived as a smell.
gene families in the human genome (14). In rodents, it has been shown that each olfactory neuron expresses only one member of the olfactory receptor genes. Neurons expressing like receptors are restricted to a regional zone oriented from a dorsal-medial to ventral-lateral orientation; however, within these regions, the expression pattern appears random among other receptors expressed in the similar area. These neurons expressing like receptors extend axons that remarkably converge together within the lamina propria to eventually converge exclusively to, on average, two glomeruli within each olfactory bulb (15). In humans, this receptor-to-glomeruli ratio is much broader and calculated to be on the order of 1:16 (16). This receptor-specific patterning is the basis of an odorant map whereby an odorant stimulates a subset of olfactory neurons that in turn activate a specific set of glomeruli in the olfactory bulb (17,18). Therefore, an odor is coded by the multiple receptor neurons it stimulates and in turn the resulting unique pattern of glomerular activation is transmitted in an unclear fashion to higher brain centers and perceived as a smell.
ETIOLOGY OF OLFACTORY LOSS
Numerous causes for a decrease in ability to smell have been described, but multiple reports from various smell and taste centers have consistently recognized similar etiologies (19). The three most common are smell loss relating to head trauma, upper respiratory infection (URI), and chronic rhinosinusitis (CRS). Another large category of smell loss is idiopathic smell loss in which no identifiable source of olfactory dysfunction can be determined. There is also a known progressive loss of sense of smell across the population related to aging with a significant drop in ability after the age of 65 and occurs in greater than one-half of people between the ages of 65 to 80 (20). The cause of this age-related loss is unclear but is independent of neurodegenerative disease. Possible theories for this loss include eventual wear-and-tear of the olfactory neurons from the cumulative effect of exposure to environmental toxins and/or progressive depletion of the regenerative capacity of epithelial basal/stem cells and genetically programmed cell death.
Smell loss related to head trauma is usually easily identified by history; however, even mild injuries to the head can result in significant olfactory disability (21). Often, in severe head trauma, the loss of smell is recognized relatively late in the recovery period due to cognitive alterations and the presence of additional separate severe injuries. The loss is thought to be a result of injury to the frontal lobes and olfactory bulbs through direct injury or commonly from a blow to the back of the head and rebound contusion to the frontal regions through a coup-countre coup injury. Another possible mechanism of traumatic smell loss is through the shearing of olfactory neurons where they transverse the cribriform plate as the brain and olfactory bulbs violently shift during trauma. Recovery from traumatic smell loss is variable but sometimes permanent with rare complete recovery and improvement in only 10% to 35% of patients (22,23,24). Usually, if return of function occurs, it generally begins within a year of the trauma.
Transient smell loss often occurs with URIs due to inflammatory swelling and obstruction of the olfactory clefts. Less commonly, a longer lasting or permanent deficit occurs after recovery from a URI. Although never confirmed, the loss is thought to be a result of a viral infection. Parainfluenza 3 has been implicated as the possible viral agent based on epidemiologic studies and molecular analysis of nasal epithelium from patients with URI-related anosmia (25,26). Even less clear is the site and mechanism of damage. Epithelial abnormalities have been found in many biopsy studies, but damage isolated to the olfactory bulbs is also possible. Women are afflicted more commonly than men in this type of smell loss, and the elderly may be more susceptible. It more often results in hyposmia instead of anosmia and the chance for recovery is relatively higher than other common forms of smell loss (27) with recovery rates between 32% and 67% (22,23).
CRS is often associated with a decreased or absent ability to smell. This can occur either with or without polyps commonly resulting from obstruction of odorant flow to the olfactory cleft. A description of fluctuating ability to smell with periods of complete return of function is rarely attributed to anything other than CRS; however, patients may also describe a gradual loss or complete anosmia years in duration. Olfaction usually returns, especially in the case of CRS with polyps, with aggressive medical treatment often including systemic steroids. However, inflammation directly involving the olfactory mucosa in patients with CRS has been demonstrated in nasal biopsies, (28,29,30) and a resulting permanent form of smell loss can occur that is eventually unresponsive to steroids. This CRS-related permanent olfactory loss has been associated with the finding of apoptotic cells within the olfactory epithelium (31) and supports the prudence of aggressive medical and/or surgical management of this disease in order to prevent longterm damage.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


