Normal Flora of the Human Conjunctiva and Eyelid
Regis P. Kowalski
Melvin I. Roat
Paul P. Thompson
Introduction
Microorganisms are present on the normal human conjunctiva and eyelid without causing ocular disease. These isolates are either contamination from the environment, spread from other areas of the body to the eye, or colonizers that thrive independently. Bacteria are the principal microbe isolated from the normal conjunctiva and eyelid. The types and numbers of bacteria are limited by human and bacterial defense mechanisms. Any deficiencies or shifts in these defenses can alter the flora. The microbial flora of the normal and compromised human conjunctiva and eyelid are discussed in this chapter.
Formation of Normal Flora
Colonization versus Recurrent Contamination
Microbes may be present on the eye as the result of two different processes. The first, colonization, results from the establishment of a stable, independent, virtually self-perpetuating community of microbes. Sometimes the colonization is characterized by biofilm formation. A biofilm is defined as a functional consortium of microorganisms organized within an extensive exopolymer matrix.1,2 The proliferation of the microbes is balanced by the host defenses, resulting in a stable number of microbes being maintained over a long period without producing disease. The second mechanism, recurrent contamination, occurs when host defenses are sufficient to eliminate the micro-organism after exposure, but the flora are continuously reintroduced from adjacent ocular structures. When recurrent contamination is frequent or continuous, it results in a stable microbe population in which, as in colonization, the microbe count can remain stable over a long period. New microbes can be introduced from the environment, adjacent tissues, or distant tissues via fomites (e.g., hand to eye).
Many of the microbes found on the eyelids have colonized the eyelids and others are introduced via continuous contamination from adjacent skin and, to a lesser extent, from the upper respiratory tract (the nose) and the upper gastrointestinal tract (the mouth). This is in contrast to the conjunctiva where the main source of microbial contamination is usually from the eyelids.
Microbe Factors
Microbial adherence is important for colonization of the eyelids, but it is especially critical for colonization of the conjunctiva where mechanisms exist to scrape and wash out the conjunctival surface. Bacteria can adhere to epithelial cells using several mechanisms. Bacterial cell wall surface glycoprotein adhesins (invasins) attach to epithelial cell membrane surface protein receptors (integrins).3 For example, the teichoic acid fraction of the Staphylococcus aureus cell wall is an adhesin to epithelial mucosal cells.3 Other microbial cell wall components (microbial surface components recognizing adhesive matrix molecules [MSCRAMM]) facilitate microbial adhesion to epithelial cell surface ligands (e.g., collagen, laminin, fibronectin, fibrinogen, vitronectin, heparin sulfate, thrombospondin and elastin).3 Some Gram-negative bacteria, such as Pseudomonas sp., Neisseria sp., Moraxella sp., and Haemophilus sp., use pili (fimbriae) to adhere to epithelial cells.3 Biofilm (slime) production can both promote adhesion and protect the bacteria from other host defenses.3
Microbes interact with other microbes in several ways. The receptor/attachment sites may be occupied by normal flora, thus limiting access to receptor/attachment sites by potential pathogens. Bacteria protect themselves from other bacteria by producing bacteriocins. These proteins are lethal to closely related bacteria. The metabolic end products of bacteria, such as acetic and lactic acids, can also inhibit the growth of other bacteria.
Host Defenses
The ocular surface is a moist, warm tissue with a high oxygen tension and an ample supply of nutrients, resulting in ideal conditions to support bacterial growth. Without adequate host defenses, contaminating microbes could colonize; colonizing microbes could increase in numbers; large numbers of microbes could overwhelm the defenses and invade; and invading microbes could infect.
The eye has several anatomic, physical, biochemical, and immunologic defense mechanisms against microbes. Anatomic and physical defense mechanisms include the following: (a) An intact epithelium helps to prevent most microbes from invading the conjunctiva and eyelid. (b) The eyebrow helps to divert liquids moving down the forehead, around the eye. (c) The eyelashes filter dust from the air and prevent contaminated particles or liquids moving along the skin from reaching the ocular surface. (d) The blink reflex, which is initiated when an object or insect touches the eyelashes, closes the eye and prevents access to the ocular surface. The normal blinking action of the eyelids has two effects: It scrapes the cornea and exposed conjunctiva, removing adherent microbes and accumulated mucus and desquamating epithelial cells, (which may be both food and habitat for bacteria). In addition, the closing action, which progresses from lateral to medial, drives the scraped material to the lacrimal puncta where it is sucked off the ocular surface into the lacrimal duct and then into the nose. (e) Normal and excessive tearing washes bacteria and debris from the conjunctiva. (f) Conjunctival and eyelid skin epithelial cells are frequently and continuously desquamated, which prevents invasion by microbes and leads to shedding of microbes adherent to the superficial layer.
Biochemical defenses include products produced by the eyelids, conjunctiva, and lacrimal glands. Some bacteria and fungi are vulnerable to unsaturated and long-chained fatty acids produced by the human meibomian glands.4,5 The lipids also produce an acidic environment that selects against some types of bacteria.3 The tear film contains many soluble components to limit microbial flora. Lysozyme is lethal to Micrococcus sp. and other bacteria (in the presence of specific immunoglobulins and complement). Metal-chelating proteins (e.g., lactoferrin) bind iron, which interferes with bacterial iron uptake and metabolism. This interference can decrease bacterial virulence and allow some bacteria to be more susceptible to other antimicrobial tear components. Lysin acts on the cytoplasmic membranes of bacteria leading to lysis.
The tears also contain complement and immunoglobulins. The components that are found in tears are the substrates for the 3 complement pathways (classical, alternate and lectin), which are cascades of reactions that result in an effective microbiological defense.6,7 Complement, when activated, leads to (a) chemotaxis of effector cells; (b) opsonization resulting in enhanced phagocytosis of bacteria and protozoa; (c) production of an anaphylatoxin; which leads to smooth muscle contraction, increased vascular permeability, mast cell degranulation, neutrophil chemotaxis and activation, and (d) lysis of virus-infected cells, bacteria, and protozoa. Immunoglobulins act against bacteria by several mechanisms. IgA (a) prevents bacteria from adhering to the conjunctival epithelium; (b) activates the alternate complement pathway; and (c) neutralizes viruses and toxins. IgG binds to microbes resulting in opsonization (independent of complement) with subsequent enhanced phagocytosis of bacteria and protozoa; antibody-dependent cell-mediated cytotoxicity of bacteria, virus-infected cells and protozoa; neutralization of viruses and protozoa; and activation of the classic complement pathway with all of the resulting effects listed above.
The cellular elements of the immune system are also active against microbes. Polymorphonuclear leukocytes can phagocytose bacteria, fungus, and protozoa. The process is facilitated by the binding of opsonizing antibodies, complement, or both. Macrophages can act as antigen-presenting cells and phagocytose bacteria and protozoa; and they can play a major role in defense against fungi, mycobacteria, and some protozoa when lymphokines are activated. Cytotoxic T cells and natural killer cells are capable of recognizing and killing virus-infected cells, independent of immunoglobulins. Natural killer cells, effector cells of antibody-dependent, cell-mediated cytotoxicity, are active against bacteria, virus-infected cells, and protozoa. Mast cells degranulate when specific IgE on their surface binds to protozoan antigens. The released effector molecules include histamine (leading to excess tearing in addition to other effects) and chemotactic factors, among others. Eosinophils degranulate when they contact IgE coated protozoa releasing effector molecules, such as major basic protein, which is highly toxic to protozoa.
Other innate immune mechanisms that contribute to ocular defense against infection include antimicrobial peptides and proteins that have demonstrated inhibitory activity to bacteria, fungi, virus, and parasites.8,9 These naturally occurring antimicrobial peptides have been shown to secrete from the ocular surface.10,11 Defensin-like chemokines and human defensin have been shown to inhibit adenovirus directly.12
The conjunctiva and eyelid have a rich blood supply, which facilitates a profuse response by the humoral and cellular elements of the immune system. The conjunctiva and eyelid also have an excellent lymphatic network and local conjunctiva-associated lymphoid tissue, both of which are important in the afferent arm of the immune response.
A balance of the bacterial and human defense mechanisms dictates the normal microbial flora of the human conjunctiva and eyelid. Infection results when an imbalance exists between proliferation of the microbes and the invasive potential of the microbes, on one hand, and the defenses by other microbes that make up the flora and the host defenses, on the other. Because an intact, stratified squamous epithelium is not easily invaded by most pathogens, the first step leading to infection is attachment of microbes to the epithelial surface. Adhesion occurs more easily to damaged than to nondamaged epithelium. Microbial attachment is not necessarily followed by epithelial cell penetration. Some microbes infect only the epithelial surface, whereas others invade the epithelial cell or subepithelial tissue.
The ocular surface and host defenses are different when the eye is closed during sleep compared with the open eye. During sleep, the ocular surface is dryer because of a lower volume of tear production (which may be due to reduced light and sensory stimulation of the lacrimal gland13), and the conjunctival vessels are dilated. A subclinical inflammatory state exists with an influx of polymorphonuclear cells,14,15,16 an increase of fibronectin17 from the blood vessels, interleukin-8 from the tears,18 and the conversion of complement C3 to C3c.14 The concentration of secretory IgA is 40 times greater in the closed eye than in the open eye.15 The closed eye also results in a decrease of oxygen to the conjunctiva and ocular surface, which alters the epithelium and opens receptor sites to bacterial attachment.19
Normal Bacterial Flora
Conjunctiva
The human conjunctiva is first exposed to microbes when an infant passes through the birth canal.20,21 Right after birth, the eye is exposed to environmental sources of bacteria (e.g., air, contaminated material, and people). The flora obtained from the birth canal is predominantly aerobic and anaerobic bacteria (Table 1). The most common aerobic bacteria isolated immediately after birth from the conjunctiva are Staphylococcus (aureus, coagulase-negative), diphtheroids (Corynebacterium sp.), Streptococcus sp., Enterococcus sp., and Escherichia coli. Propionibacterium acnes, Bifidobacterium sp., and Bacteroides sp. are isolated anaerobically from the newborn conjunctiva after vaginal delivery. Lactobacillus sp., a microaerophilic bacteria, can also contaminate the conjunctiva from the vaginal tract. Other anaerobes and aerobes have been isolated less frequently (Table 1). The conjunctiva is sterile in 20% to 87% of infants born vaginally.22 The conjunctiva cultured after cesarean section is sterile in 80% to 95% of infants.22,23 If cesarean section is delayed for more than 3 hours after the sac is ruptured, conjunctiva will be sterile in 45% of newborns. Eder et al.,24 in a recent study from Argentina and Paraguay, also reported a culture-positive rate higher in vaginal births than in cesarean section deliveries.24
During the first several days of life, the infant’s own flora in skin, respiratory tract, and gastrointestinal tract, and the flora of the surrounding environment are important new sources of bacteria for the conjunctiva. After birth, before prophylaxis, a dramatic increase occurs in the frequency of culture-positive conjunctivae, from as low as 13% of newborns to 98% of 3- to 5-day-old infants. Bacteria isolated include Staphylococcus sp., Streptococcus sp., or Moraxella (Branhamella) catarrhalis.22
The literature (Table 2-41.1) reports that from birth to old age, the conjunctiva is culture-negative for bacteria in 39% (range 73% to 14%) of cases. The frequency of isolating bacteria from the conjunctiva increases with age (Fig. 2-41.1). Coagulase-negative Staphylococcus, diphtheroids (Corynebacterium sp.), Propionibacterium acnes, and Staphylococcus aureus are the most frequently isolated bacteria from the normal conjunctiva. In general, the density of isolation is limited to less than 10 colonies per culture compared with the confluent growth seen with an infection. Streptococcus sp. are isolated more commonly from children than adults. Propionibacterium acnes, diphtheroids, and Gram-negative bacteria are more frequently found in adults25,26. Less commonly, other Gram-positive and Gram-negative bacteria are isolated from the conjunctiva (Table 2-41.2). These isolates are probably transitory residents passed to the eye from other parts of the body and the environment via the adjacent skin or by the hands.
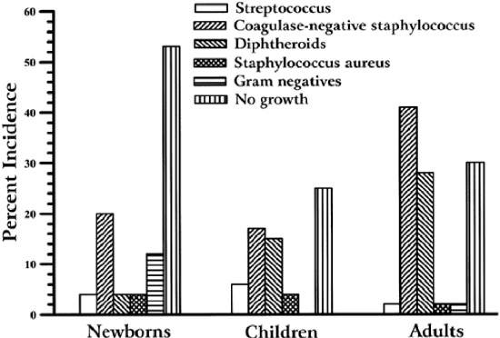 |
Table 2-41.1. Bacterial Flora of Normal Human Conjunctiva from Birth through Various Age Groups Number of isolates (%) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Table 2-41.2. Normal Flora of the Normal Human Conjunctiva Before Prophylaxis and Surgery Number of isolates (&) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


