Chapter 19 Mechanisms of Normal Retinal Adhesion
Independent of its clinical relevance, the study of retinal adhesion is also of considerable physiologic interest. No anatomic junctions bridge the mammalian subretinal space,1 yet this primordial dural cavity remains collapsed and, indeed, tightly closed throughout a lifetime of ocular movement and tugging from the vitreous. Our current knowledge of this system is still incomplete, but evidence suggests that adhesion depends on a mixture of anatomic, physical, and metabolic factors.2 This chapter reviews these factors and considers clinical and therapeutic implications.
Models for measuring retinal adhesion
In vitro methods
Adhesive force falls rapidly after enucleation or death,3,4 as is discussed later, and this puts a constraint of time on techniques for measuring adhesiveness in vitro.5,6 One approach is to peel the retina within a fluid bath while recording the required force with a transducer.3 A faster method is to measure the amount of RPE pigment that remains attached to the retina after separation as the index of adhesiveness.7 An eye can be enucleated and small strips of the eyecup prepared within 30 seconds, after which the retina is gently peeled by hand from the RPE. The stronger the adhesive force, the more pigment adheres to the peeled retina.
In vivo methods
Kita et al.8 developed the most direct technique for quantifying adhesiveness within the living eye. A small detachment is made in the eye by injecting fluid into the subretinal space through a micropipette, and a second micropipette is inserted simultaneously into the detachment cavity to measure fluid pressure. The pressure that is required to expand the detachment can be converted mathematically, by Laplace’s law, into a value for adhesive force at the margin of the detachment (where the separation is taking place). This technique allows the evaluation in living animals of the effects of drugs and other agents that modify retinal adhesion.9,10
Adhesive force and environmental factors
Magnitude of adhesive force
In vitro measurements, 5–20 minutes after enucleation, have shown that about 25 mg of force is required to peel a 5-mm strip of rabbit retina from the RPE.3,11,12 Pressure measurements in living eyes have shown an adhesive force in the rabbit of 100–180 dyn/cm.8,9 Adhesion is stronger in cats and monkeys, which show mean values for adhesive force that are 180% (for cats) and 140% (for monkeys) of that in the rabbits.9
Sensitivity to temperature and ionic environment
Retinal adhesiveness drops rapidly postmortem at 37 °C, but remains near control levels for hours at 4 °C7,13–15 (Fig. 19.1A). These effects of temperature are reversible. Figure 19.1B shows that changing the temperature from 37 °C to 4 °C or vice versa causes retinal adherence in the rabbit16 to rise or fall, respectively, and repeatedly. This reversibility has also been documented in primate and human tissue.14,15,17 The mechanism of these reversible temperature effects is still obscure. Temperature could act directly on physicochemical components of adhesion or modulate metabolic systems. Some of the adhesion-enhancing effect of cold temperature may result from sodium pump inhibition and secondary tissue swelling, which would make the interdigitated outer segments and RPE microvilli harder to separate.3,13 Because cellular swelling is a pathologic event, the actions of cold temperature may not be entirely relevant to normal adhesive processes.
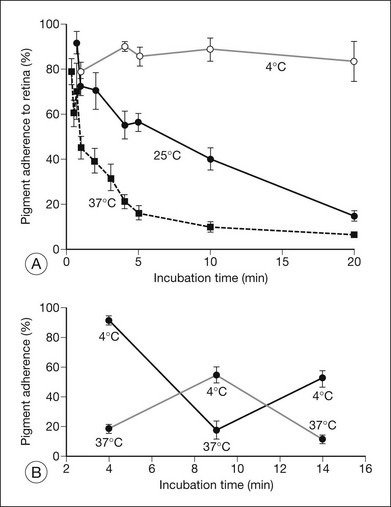
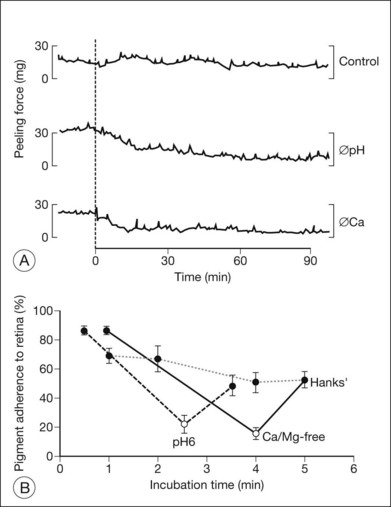
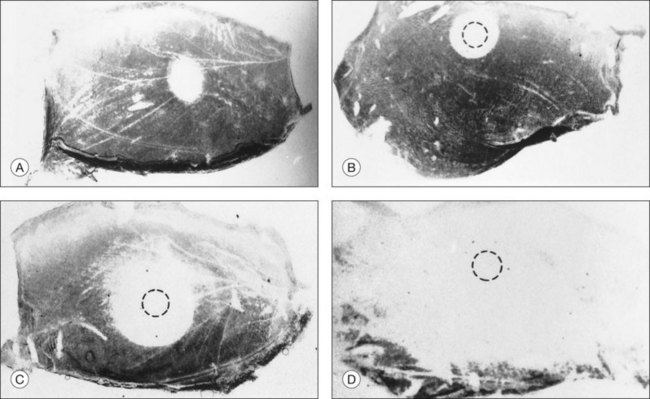
The ionic environment appears to be critical to adhesive strength. In in vitro experiments using tissue from rabbits, humans, and other primates, lowering the pH from 7.4 to 5.5, or removing calcium and magnesium ions from the bathing solution, weakened adhesive force and accelerated the rate at which adhesion falls postmortem11,13,15,17 (Fig. 19.2A). These changes can be rapidly reversible16 (Fig. 19.2B), although cold temperature will block or mask the effects of pH and calcium ions. In vivo, the removal of calcium ions from the subretinal space weakens retinal adhesion in rabbits to about 30% of normal.18 In other words, calcium appears to be a necessary element for the maintenance of normal adhesiveness in the living eye.
Fluid pressure: hydrostatic and osmotic
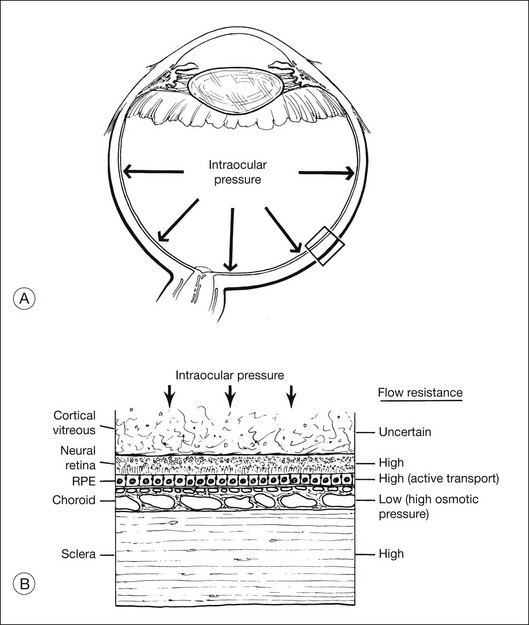
Fluid is driven passively from vitreous to choroid by both intraocular pressure and the osmotic pressure of the extracellular fluid in the choroid (estimated to be about 12 mmHg in the rabbit,19 but possibly lower in humans20). However, under normal conditions there appears to be relatively little posterior flow, and most of the fluid that enters the eye as aqueous leaves the eye through anterior drainage channels.19,21 The posterior route is limited because the retina and RPE22,23 provide a substantial resistance to water movement, and little fluid can cross these layers under the available heads of pressure. A side-effect of this retinal flow resistance is that the outward movement of fluid acts to push the retina against the RPE (Fig. 19.3). This is one mechanism contributing to normal retinal attachment. The actual pressure difference across retina or RPE is probably small because intraocular pressure is ultimately contained by the sclera, and tissue pressures will therefore equalize to a large degree between the vitreous and choroid. However, Fatt and Shantinath22 have calculated that even a very small pressure difference (only 0.52 × 10−3 mmHg) across the retina would generate a force sufficient to keep the retina firmly fixed against the wall of the eye. This low value may explain the fact that no one14,19 has been able to measure a pressure difference between the vitreous cavity and subretinal space.
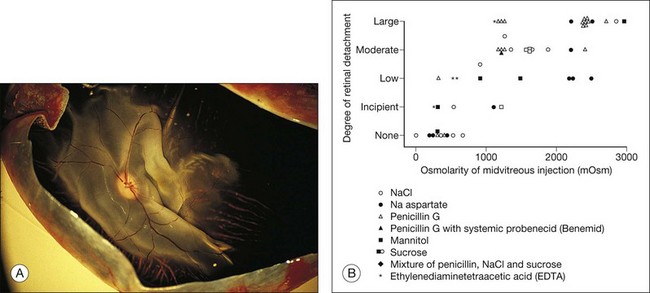
These physical forces can also work in the other direction and cause retinal detachment. For example, if hyperosmotic fluid is introduced into the vitreous cavity, fluid moving from the choroid into the vitreous will elevate the retina24,25 (Fig. 19.4). This is an important concern in the evaluation or use of intravitreal drugs that may damage the retina osmotically independently of pharmacologic effects.
It may seem puzzling that, although most intraocular fluid leaves the eye anteriorly rather than posteriorly, ample animal data26–30 indicated that the RPE can pump fluid from the subretinal space to choroid at a very high rate (about 0.3 µL/h/mm2 of RPE) comparable with that of aqueous secretion. How can the RPE have such enormous power for fluid removal, yet very little fluid leaves the normal eye by a posterior route? The answer probably lies in the flow resistance of the retina. Under normal conditions, given the flow resistance of the tissue and the tiny pressure differential across it,22,23 water will cross the retina only at a very slow rate. In other words, the tissue resistance is rate-limiting, and most aqueous fluid must leave by the anterior route. Under pathologic conditions of retinal detachment (whether rhegmatogenous or nonrhegmatogenous), fluid is present within the subretinal space and is thus available for transport at a maximal rate. Measurements in humans for the resorption of detachments showed a rate of 0.11 µL/h/mm2 of RPE, and the rate may well be higher in eyes without pathology.31 This translates into roughly 3.5 mL of fluid per day, which explains why a rhegmatogenous detachment can settle within 24 hours, and emphasizes the power of RPE fluid transport.
The clinical importance of fluid pressure as a factor in preventing retinal detachment is uncertain. It undoubtedly helps to keep the retina in place when the retina is intact, but raising intraocular pressure in rabbits to 38 mmHg or lowering it to 0 mmHg had only a modest effect on the rate of subretinal fluid absorption.32 Furthermore, clinical retinal (as opposed to choroidal) detachment is uncommon in hypotonus eyes, and even a small hole in the retina would theoretically allow fluid to reach the subretinal space and circumvent fluid pressure mechanisms of adhesion. Retinal holes are, in fact, found in about 10% of autopsy eyes without detachment,33 but many of these holes may not be functionally open because of surrounding pigmentation or because of “tamponade” by the cortical vitreous gel.34
Once the retina has detached and adhesion mechanisms that depend on retina–RPE contact can no longer work, fluid dynamics become much more important, even in the presence of a hole. Hammer35 has calculated that fluid entering a retinal hole over a scleral buckle will, by its flow pattern, create a suction force that pulls the retina backward against the buckle.
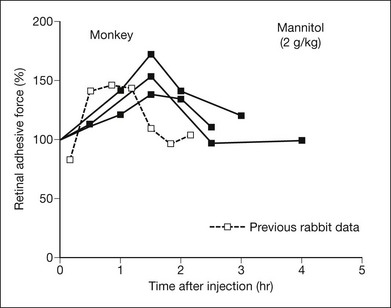
Osmotic pressure can be manipulated more easily and may turn out to have therapeutic applications. Both in vitro and in vivo experiments, using rabbits and primates,9,36,37 have shown that an intravenous injection of mannitol will increase retinal adhesiveness by roughly 50% within 1–2 hours of injection (Fig. 19.5). The magnitude of the effect correlates rather closely with blood osmolality. Mannitol is known to enhance the absorption of fluid out of the subretinal space.38 Some of the adhesive effect may come from fluid absorption “pulling” the retina against the RPE; however, most of the effect probably comes from dehydration of the subretinal space (which enhances the binding properties of the interphotoreceptor matrix (IPM)), because the mannitol effect is still evident in excised tissue where there is no flow toward the choriocapillaris.
Vitreous support and other physical aspects of adhesion
The role of the vitreous in creating retinal detachments through traction and contraction is well known. The role of the vitreous in maintaining retinal attachment is less clear. Vitreous gel has a physical structure that may help to keep the retina in place,34,39 although retinas do not just come off when vitreous detachment or syneresis occurs. A thin cortical layer of vitreous might remain after vitreous detachment or syneresis and serve as a seal or tamponade for retinal holes,34 thus aiding the action of fluid pressure in keeping the retina apposed (Fig. 19.3).
There is other, indirect, evidence for a vitreous role in adhesion. It is very difficult to produce and maintain an experimental rhegmatogenous detachment if the vitreous body remains intact.24,40 Vitreous removal (either mechanical or with hyaluronidase) presumably weakens the tamponade and the structural qualities of the gel, but it also allows liquid vitreous to reach the subretinal space. The status of the gel may help account for the vastly different incidence of retinal detachment among young individuals, in whom the gel is largely intact, and among older individuals, in whom syneresis and vitreous detachment have occurred. A retinal hole in a young eye is blocked by gel pressure and can seal uneventfully; a hole in an old eye is more likely to allow fluid to enter the subretinal space and cause detachment. On the other hand, the integrity of the vitreous gel seems of little consequence to the formation or persistence of experimental nonrhegmatogenous detachments32 or to the retinal adhesive force measured by peeling,11 so the role of the vitreous gel in attachment may be more one of preventing pathologic fluid access to the subretinal space than one of providing a direct adhesive support or force.
The weight of retina is potentially a factor in attachment and detachment, as influenced by gravity and ocular movement. Once a detachment has occurred, the influence of gravity is well known, because appropriate positioning of the patient can be enormously effective in inducing retina to settle. However, it is hard to envision retinal weight as a major factor in retinal attachment under normal conditions,27,32 because our upright posture and eye movements would dictate that benefits and adverse effects be simultaneously present in different parts of the eye.
Mechanical forces inside the subretinal space
Anatomic bridges do not exist between retina and RPE,1 but it is reasonable to ask to what extent matrix material between the layers serves as “glue” and to what extent the physical interdigitation of photoreceptor outer segments and RPE microvilli serves to hold the tissues together.
Mechanical interdigitation
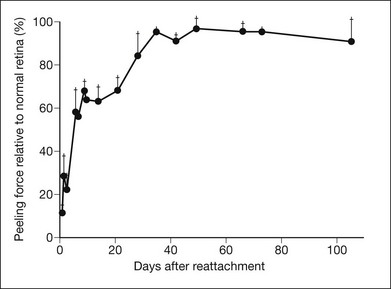
RPE microvilli wrap closely around the tips of the outer segments (Fig. 19.6), and this connection is strong enough to allow for daily phagocytosis of outer-segment fragments as the photoreceptors renew their disc material.1,41,42 However, as a factor in adhesion, interdigitation seems of variable importance. The RPE microvilli are subject to continuous cellular remodeling during the outer-segment renewal cycle. After surgical repair of a retinal detachment, reattachment takes place rather promptly, before the outer segments have regenerated and microvillous connections have been re-established.43,44 Interdigitation begins to develop within 3 days of reattachment, but retinal adhesiveness does not return to normal until 5–6 weeks after reapposition of experimentally detached retina in the rabbit45 (Fig. 19.7).
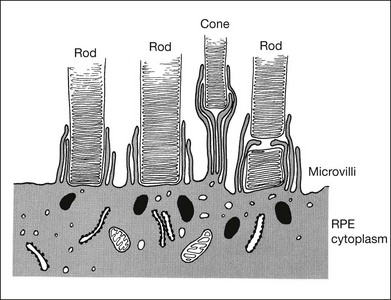
The mechanisms by which interdigitation could produce adhesion are not known. During outer-segment phagocytosis, the microvilli indent the outer segments and must physically impede attempts to withdraw them. Close ensheathment might also provide a frictional resistance to withdrawal, just as a finger is hard to pull from a narrow tube. There may be electrostatic forces that oppose separation of the membranes.46 The magnitude of all of these effects will also depend on other factors, such as the composition of the intervening matrix and capability of the RPE microvilli for motility and remodeling.
Interphotoreceptor matrix properties
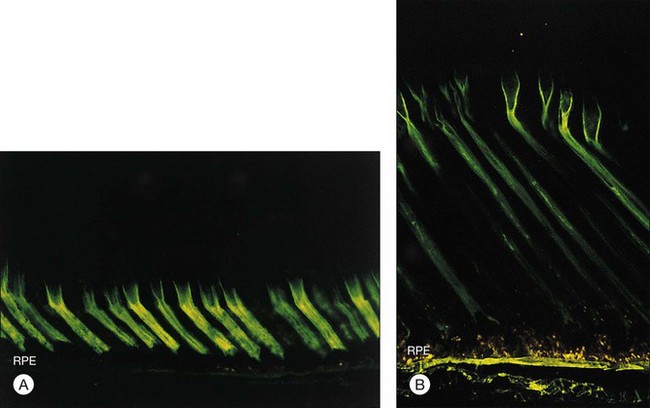
The IPM is a viscous material composed largely of proteins, glycoproteins, and proteoglycans47,48 but containing a substantial concentration of glycosaminoglycans.49 The idea that IPM might serve as a viscous glue was proposed many years ago by Berman,50 but current evidence indicates the IPM is not just a layer of glue but has component structures that play a role in the adhesive process. For example, the cones are surrounded by a specialized sheath of matrix51,52 that can be recognized histochemically by the binding of peanut agglutinin (PNA) (Fig. 19.8A). These cone matrix sheaths remain attached to both RPE and photoreceptor cells when the neural retina is peeled from the RPE.48,51,53 If fresh primate or human retina is peeled from the RPE, the matrix material will stretch markedly before breaking (Fig. 19.8B), suggesting a rather strong bonding at both the outer-segment and RPE interfaces.17,54 The evidence for a bonding function of the IPM is strongest with respect to the cone matrix sheaths, but the structural material that surrounds the rods may play a similar role. It can be recognized histochemically by staining with wheatgerm agglutinin (WGA) or other lectins,55,56 and stretching of WGA-staining material may be observed on peeling freshly enucleated eyecup material.17,54,57 The role of newly recognized matrix components such as galectins and IPM proteoglycans remains to be determined.58,59
The ability of cone or rod matrix sheaths to serve as a structural bond between retina and RPE may well depend on the presence of specific receptors that bind IPM components to the cell membranes. (“Cell adhesion molecules” represent a group of substances that mediate adhesion between individual cells or between cells and substrates.60) Experiments have shown that extracted IPM material does not automatically stick cells to one another,61 but adhesion is unlikely unless specific receptors are present to bind the macromolecular substances. Several receptor systems, including sites for fibronectin, integrins, and mannose, have been proposed and disputed as being involved with retinal adhesion.48,62–65 There is functional evidence that inhibition of chondroitin sulfate proteoglycan synthesis with xylosides causes spontaneous detachments in primates.48,66
Other types of evidence support the concept that IPM contributes significantly to retinal adhesion. Physical factors that affect adhesiveness, such as temperature, pH, and calcium concentration, are known to be modulators of the physicochemical properties of IPM macromolecules and may also affect the bonding of large-molecular species at receptoral sites.67,68 There is also a marked loss of retinal adhesiveness when the subretinal space is exposed to enzymes that degrade matrix components,15,69 such as chondroitinase ABC (which degrades chondroitin sulfate70) or neuraminidase (which breaks sialic acid bonds71). These enzymes, given intravitreally or into the subretinal space itself, weaken adhesiveness markedly in correlation with cytochemical evidence of damage to the IPM (Figs 19.9 and 19.10). It is unknown whether the effects of these enzymes on adhesiveness are a result of changes in viscosity or damage to the structural elements of the IPM. However, adhesiveness returns to normal roughly 3 weeks after exposure of an eye to neuraminidase or chondroitinase ABC, and the recovery of adhesiveness correlates closely with the recovery of normal PNA-binding properties in the IPM.72
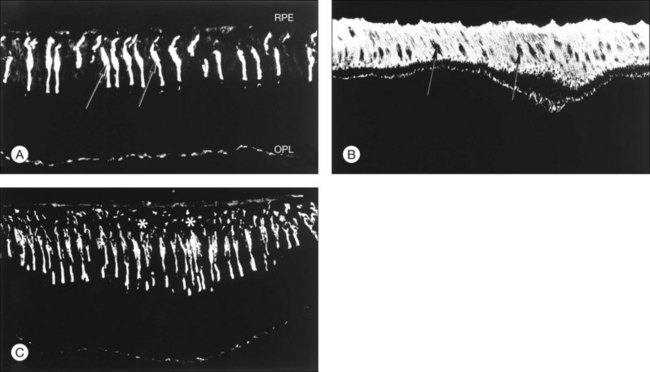
Experiments have shown that the chemical nature of the IPM may change with light and dark adaptation.73 To the extent that photic effects might modify the viscosity or bonding properties of the IPM, light could be a modulating factor of retinal adhesion. However, experimental data on adhesion in the light and dark have been contradictory and of uncertain significance.11,74,75
Subcellular components and mobility
Subcellular components of the RPE, such as microtubules and microfilaments, will be relevant to retinal adhesion to the extent that they influence remodeling and movement of the RPE microvilli.76 Actin filaments control melanin movement in the amphibian RPE11 and are present in mammalian RPE,77,78 but a movement of melanin granules has never been observed in mammals. A few attempts have been made to alter adhesion by inhibiting subcellular components of the RPE. Cytochalasin B, which blocks microfilaments, was found to have no effect on adhesion when the drug was presented to excised tissue briefly after enucleation.12 However, injecting cytochalasin into the vitreous 4–72 hours before enucleation caused a 70–90% reduction in pigment adherence as a measure of adhesiveness.79
When retina detaches, the surface morphology of the RPE changes literally within minutes.23 The evolution of these changes can be altered by both colchicine (which dissembles microtubules) and cytochalasin,80 suggesting that microtubules and microfilaments are important to apical RPE morphology. Actin filament contraction and subcellular motility are known to require calcium,81 the absence of which is severely detrimental to retinal adhesion. However, we do not know whether the action of calcium in weakening adhesiveness relates to this mechanism or to effects that calcium may have on transport systems and matrix-binding properties.
Critical dependence on oxygen
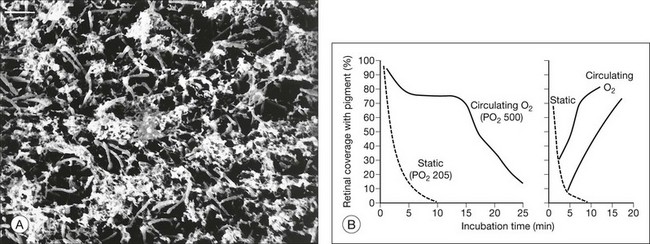
It was noted previously that adhesiveness in living rabbit eyes falls severely within a few minutes of death5 (Fig. 19.1A). Furthermore, just 1 minute of ocular ischemia in living rabbits weakens adhesion dramatically, whereas restoration of circulation restores adhesiveness.82,83 The postmortem adhesive loss appears to be slower in primates and humans, and under similar in vitro conditions, at least a moderate adhesive force was measurable for 30 minutes or more after enucleation of primate and human tissue15,17,83 (Fig. 19.11A).
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


