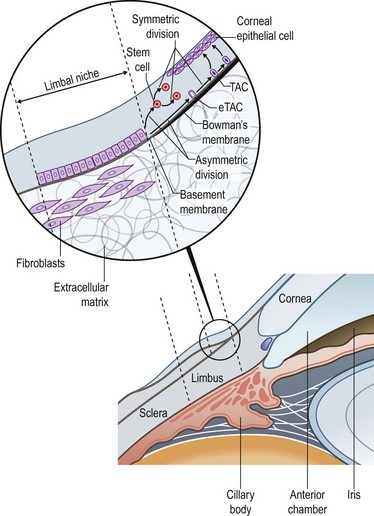
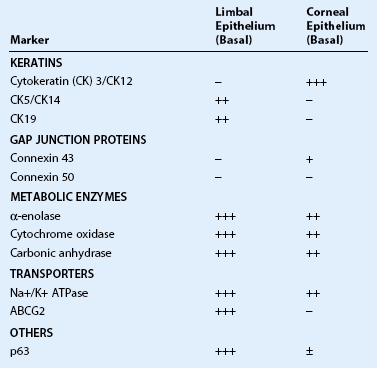
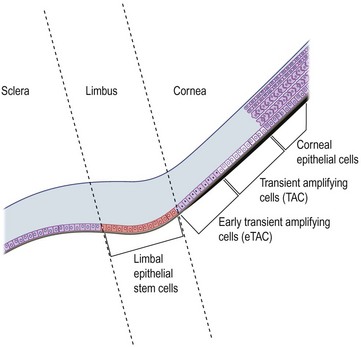
5 The narrow transitional zone between the corneal and bulbar conjunctival epithelium represents the limbal epithelium. However, due to the lack of distinct borders, there are various anatomic definitions of the limbus as defined by anatomists, pathologists, histologists, and surgeons. The most accepted definition delineates the inferior border of the limbus as a line between the outer border of Bowman’s layer and Descemet’s membrane, and the exterior border as the start of scleral collagen fibers and outside border of the Schlemm’s canal, 1.5 to 2 mm outside the inferior border (Fig. 5.1 ).1 This region has an important barrier function and prevents conjunctival overgrowth onto the cornea. Histologically, the non-keratinized stratified limbal epithelium can be differentiated from the conjunctival epithelium, in that it lacks goblet cells. Compared to the corneal epithelium, while the superficial epithelial layers are rather similar, the limbal epithelium contains cell layers, a large number of mature (activated) and immature epithelial dendritic cells, T lymphocytes, highly pigmented melanocytes, and subjacent blood vessels. Moreover, the basal limbal epithelial cells are unique in that they are the least differentiated cells of the ocular surface epithelium.2 These cells are smaller, less columnar and have more cytoplasmic organelles. A growing body of evidence over the past years supports the theory that these cells are limbal epithelial stem cells (LESC), giving rise to the more differentiated corneal epithelium.2 Limbal epithelial stem cells reside in the limbal niche,3 where subepithelial papillae-like structures known as palisades of Vogt are seen clinically.4 The palisades of Vogt appear as radial linear structures of about 1 mm in length as observed by slit-lamp microscopy and in vivo confocal microscopy.5 This anatomical landmark provides the homeostatic microenvironment that promotes the maintenance of limbal epithelial stem cells (LESCs) in an undifferentiated state. Currently, no single marker can be used to identify LESCs definitively, which lack terminal differentiation markers. However, LESCs can be differentiated from the corneal epithelium by several markers, including p63, vimentin, α9β1 integrin, cytokeratin (CK)19, CK5, CK14, cadherin 342, and the ATP-binding cassette subfamily G member 2 (ABCG2) transporter protein (Table 5.1). Further, LESCs lack CK3 and CK12, which are characteristic for the corneal epithelium. They are heavily pigmented in order to be protected form ultraviolet light damage. LESCs produce several metabolic enzymes and proteins at higher levels than corneal epithelial cells, such as α-enolase, cytochrome oxidase, Na+-K+ ATPase, carbonic anhydrase, and glucose transporter. The functional relevance of these enzymes and proteins are yet to be elucidated. Although LESCs are slowly cycling and divide only occasionally, they have high proliferative and self-renewal capacity.3 Due to their slow cell cycling, they have a higher retention of DNA precursor analogs. However, in the event of injury, LESCs begin rapid proliferation. In order to retain a constant stem cell pool, LESCs undergo two types of cell division: a symmetric and asymmetric division (Fig. 5.1). During symmetric division, either two identical stem cells or alternatively two identical differentiated daughter cells emerge. In contrast, asymmetric division of LESCs results into a stem cell and an early transient amplifying cell (eTAC).6 These eTACs further divide and give rise to additional TACs (Fig. 5.2). TACs finally migrate centripetally towards the corneal center, ultimately forming the terminally differentiated corneal epithelial cells. This terminal differentiation of TACs into corneal epithelial cells is accompanied by specific morphological and biochemical alterations.
Limbus and Corneal Epithelium
Limbal Epithelium
Limbal Epithelial Stem Cells
Differentiation of Limbal Epithelial Stem Cells to Corneal Epithelium
Limbus and Corneal Epithelium