Iris
Cecilia S. Jung
Hans E. Grossniklaus
Amy K. Hutchinson
The iris functions as a diaphragm within the eye, separating the anterior and posterior chambers of the eye. The structure is named after Iris, the Greek goddess of the rainbow, and provides us with our characteristic eye color. Its central opening, the pupil, is the aperture that regulates the amount of light entering the eye. Although seemingly simple in structure, iris function is complex and affects image clarity. Normal and pathologic states of the efferent and afferent visual pathways are clinically evaluated via pupillary function. The iris is the most anterior part of the uveal tract and can be involved in many ocular and systemic diseases. This chapter discusses the embryology, gross anatomy, histology, and ultrastructural features of the iris.
EMBRYOLOGY
Historically, the embryology of the iris was based on the classical theory of development, which described three germ layers in the developing embryo: endoderm, mesoderm, and ectoderm.1,2,3 It is now known that the majority of the ocular mesenchyme is derived from neural crest cells.4,5,6,7 Neural crest cells are defined as those neuroectodermal cells that proliferate from the crest of the neural folds before the formation of the neural tube. Recently, certain genetic transcription factors, such as PAX6, have been shown to play a key role in orchestrating the formation of the developing tissues in the anterior segment, and mutations of such genes have been implicated in certain diseases of the iris, including aniridia.8,9
Embryologically, the structure of the iris can be divided into two different developmental layers: iris pigment epithelium (IPE) of neuroectodermal origin and iris stroma of neural crest origin. The iris stromal blood vessels begin as penetrating branches from the developing vascular network of the ciliary body (Fig. 11.1). Although data regarding the origin of both iris muscles (sphincter and dilator) are incomplete, they likely arise from neuroectoderm.4,6
The embryologic precursors of the iris are first visible at 6 weeks’ gestational age. The neural ectodermal derivatives (iris epithelium and muscle) arise from the anterior rim of the optic cup. At about 6 weeks’ gestational age, these cells begin to proliferate rapidly and extend anteriorly10 During development, an apparent dilation, called the marginal sinus of von Szily, can be seen at the tip of the optic cup between the two layers of epithelium. Although this sinus is now thought to represent an artifact, the two layers of neuroepithelium become functionally divided into the anterior and posterior pigment epithelium (Fig. 11.2).11 Embryologically, the anterior epithelium becomes pigmented while still part of the optic cup; however, the pigmentation of the posterior epithelium (derived from the inner, unpigmented layer of the optic cup) occurs later, proceeding toward the ciliary region, and is completed by 7 months of gestation.12
Differentiation of the iris stromal elements begins slightly earlier than iris neuroepithelial growth and proceeds as a wave of mesenchymal migration at about 6 weeks’ gestational age. This forms the anlage of iris stroma, a continuous pupillary membrane, and the tunica vasculosa lentis (a transient vascular network that supplies the developing lens). Iris stromal pigmentation does not occur until after 24 weeks. The developing iris lacks a pupil because the original stroma is a continuous sheet of tissue with epithelial cells growing inward, toward the center of the iris.4,11
The vascular framework of the iris also begins at 6 weeks as a blind axial outgrowth from the annular vessels in the mesenchyme associated with the rim of the optic cup and forms the anterior part of the tunica vasculosa lentis (lamina iridopupillaris). By the third month, branches of the nasal and the temporal long posterior ciliary arteries unite with peripheral vessels of the lamina iridopupillaris to form the major arterial circle. The pupil is created by cellular and vascular remodeling of the iridopupillary membrane during the fifth month of gestation.4,11
The development of the iris sphincter muscle first appears as basal infoldings in the cells of the anterior layer of neuroepithelium and can be seen by light microscopy at about 11 weeks. By the fifth month, myofibrils begin to be synthesized by the developing tissue; the muscle comes to lie free in the posterior mesenchymal layer by the eighth month of gestation. The formation of the dilator muscle fibers occurs later and is first identified in the sixth month as fine fibrils in the anterior iris epithelium.
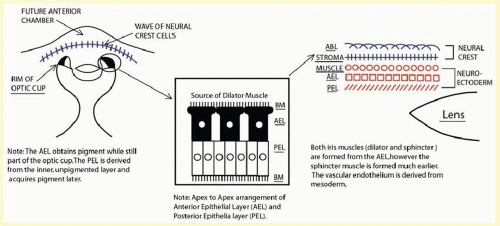 FIG. 11.1 As demonstrated, the iris is derived from both neural crest cells and the optic cup. The AEL and posterior epithelial layer (PEL) are derived from the optic cup and are oriented in an apex-to-apex pattern with BMs. When the five layers of the iris are seen in cross section, one can see that the ABL and stroma (colored in blue) are derived from the neural crest cells and the iris muscles; AEL and PEL (shaded in orange) come from the optic cup. |
The iris is near its adult size at birth. The most dynamic changes occur in the first few postnatal years and involve the color of the iris and the extracellular matrix. Embryologically, the anterior epithelium becomes pigmented while still part of the optic cup; however, the pigmentation of the posterior epithelium (derived from the inner, unpigmented layer of the optic cup) occurs later, proceeding toward the ciliary region, and is completed by 7 months’ of gestation. Postnatal changes in iris color occur by the gradual accumulation of melanin in the stroma and anterior border layer (ABL).
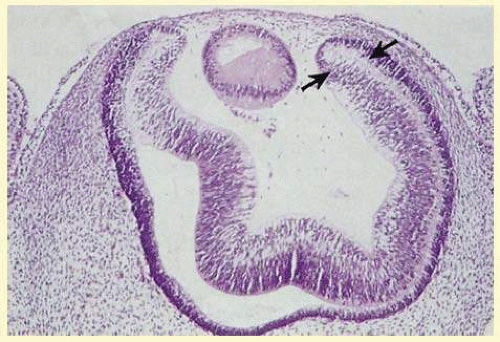 FIG. 11.2 The two layers of the neuroepithelium of the optic cup (arrows) become the anterior and posterior IPE, and the iris stroma is derived from neural crest tissue (hematoxylin-eosin, X63). |
GROSS ANATOMY
The iris measures approximately 12 mm in diameter and about 37 to 38 mm in circumference. It is thickest at its central portion (the collarette) and thinnest near the ciliary body’s insertion point (the root, the site most prone to tears from trauma). The pupil is decentered somewhat nasally and inferiorly to the central cornea with the temporal iris being broader than the nasal iris. Anteriorly, the iris is bathed by aqueous humor. Posteriorly, the central portion of the iris abuts the lens, which displaces the iris somewhat anteriorly and results in its characteristic conical configuration. Physical analyses of the factors that determine the contour of the iris are available.13,14 Peripherally, the posterior iris is bathed by the aqueous in the posterior chamber.
Using advances in anterior segment imaging, several recent studies suggest that greater iris curvature, area, and thickness are associated with narrow angles. A thicker peripheral iris could crowd angles in predisposed eyes, such as hyperopic eyes.15,16,17
The iris color is determined by four main factors: (1) the density of pigment granules in the IPE, (2) the concentration of pigment in the iris stromal melanocytes, (3) the chemical character of the melanin, and (4) the light scattering and absorptive properties of extracellular components of the iris. In contrast to what was previously believed, the number of melanocytes is relatively constant in irides of different colors. Melanosomes vary in grades depending on their maturity. Darker irides have more pigment granules, larger and higher grade melanosomes and a higher ratio of eumelanin to pheomelanin when compared with blue irides. Longer wavelengths of light are absorbed by the IPE, whereas shorter wavelengths are reflected and undergo Tyndall scattering by the stroma. In brown irides, this short wavelength light is absorbed by the melanin contained in the stromal melanocytes, but in eyes that are less pigmented, the scattering gives rise to a Tyndall blue structural color that varies with external lighting conditions.18,19,20,21 The characteristic pink color of tyrosinase-negative albino irides is predominantly driven by the transillumination red reflex and blood within the iris vessels.22
More recently, the complex genetic components of iris pigmentations are beginning to be understood. The iris color is likely inherited as a polygenic trait despite previously taught Mendelian inheritance, but this is yet to be further studied.23,24 A number of pigment genes such as OCA2, TYRP1, MAPT, and MYO5A have been isolated and implicated in the iris color.24,25
The recent use of topical prostaglandin therapy in the treatment of glaucoma has shown that the color of the iris may also change in some adult patients, which may provide insight into the possible cellular pathways that control iris melanocytes. There are several proposed explanations: (1) the drugs may cause the melanocytes to proliferate; (2) prostaglandins may act directly or through a second messenger on melanocytes, causing either increased tyrosinase activity or decreased breakdown of melanosomes; (3) prostaglandins may cause migration of stromal melanocytes; or (4) these drugs may influence the responsiveness of melanocytes to neuronal stimuli.26,27,28 More recent studies show that susceptibilities to the iris color change may also be related to genetics, basal iris color, or age.29
ANTERIOR IRIS SURFACE
The collarette of the iris is located about 1.5 mm from the pupillary margin, represents the thickest portion of the iris (0.6 mm), and divides the iris into two zones: the pupillary zone and the ciliary zone (Fig. 11.3). The pupillary zone begins at the margin of the pupil, known as the pupillary ruff, which represents the anterior termination of the IPE. Numerous crenations are seen in this area and vary with pupillary size. Thick branching trabeculae are most prominently seen in the pupillary and collarette regions. These bands are radially oriented and enclose depressions in the iris surface. Fuchs crypts are adjacent to the collarette on both its pupillary and ciliary sides. These crypts are focal defects in the anterior layer of the iris stroma, which arise from agenesis, or more likely, atrophy of this layer coincident with resorption of the embryonic pupillary membrane.19 The anterior surface of the ciliary portion has been divided into three areas: an inner smooth area, a middle furrowed area, and a marginal cribiform area (which is visible only by gonioscopy). The entire ciliary region is characterized by a pattern of radial ridges formed by underlying stromal blood vessels.20
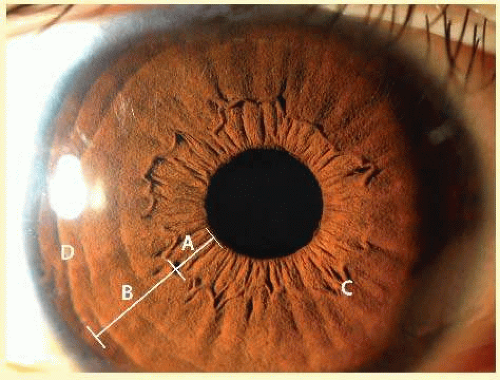 FIG. 11.3 The anterior iris surface features the inner pupillary zone (A) and outer ciliary zone (B) divided by collarette. Fuchs crypts (C) and marginal furrows (D) are present. |
POSTERIOR IRIS SURFACE
The posterior surface of the iris is much smoother and more uniform than its anterior counterpart (Fig. 11.4). The posterior surface of the iris contains two layers of epithelial cells: the anterior and posterior iris epithelia. The surface is smooth but broken by relatively shallow furrows. Two types of radial folds are identified: (1) contraction folds of Schwalbe, which are small sulci that run for a distance of 1 mm, bending over the pupillary aperture and forming the crenations on its anterior surface and (2) structural folds of Schwalbe, which begin about 1.5 mm from the pupillary border and extend between the ciliary processes. Circular contraction furrows are also present.20
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


