Mediators of Ocular Inflammation
Matthew J. Chapin
Paula J. Wun
Mark B. Abelson
Mediators are biologically active chemical compounds contained within inflammatory cells. On release from the cell, mediators act in a specific manner and at a specific site to induce a component of the inflammatory or immunologic process. Our understanding of the roles individual mediators play in the inflammatory and allergic disease has increased rapidly. This understanding allows us to identify rational and more effective means of therapy.
The mast cell is the principal effector cell for allergic reactions in the eye and other organ systems. The mast cell contains more than 30 preformed mediators, and mast cell degranulation stimulates the synthesis of many more. Mediators that are preformed and released by the mast cell, or are synthesized after mast cell activation, will be the primary focus of this review. However, other mediators with a role in ocular inflammation will also be discussed.
The mast cell surface has as many as 500,000 immunoglobulin-E (IgE) receptors, 10% of which are occupied in vivo.1 The Fc portion of the IgE molecule, the portion attached to the mast cell membrane, changes as a result of IgE cross-linking with the offending allergen, activating a serine esterase.2 This leads to an intracellular biochemical cascade causing mast cell degranulation and the subsequent release of preformed mediators, including histamine, eosinophil chemotactic factor of anaphylaxis, high-molecular-weight neutrophil chemotactic factor, and platelet-activating factor (Fig. 1). These mediators attract eosinophils and neutrophils, which restore homeostasis to the tissue. The signs and symptoms of an acute allergic reaction result from this intricate network of mediator interaction.
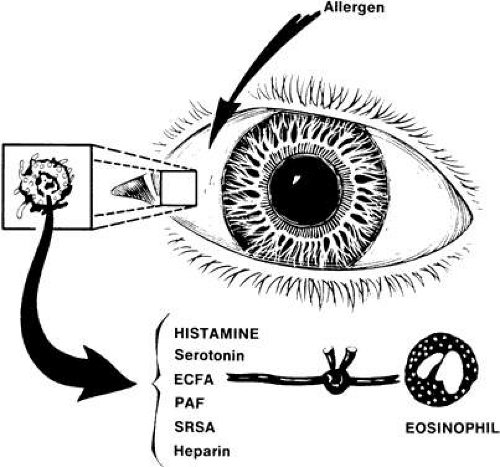 Fig. 1. Release of mast cell mediators in response to antigenic stimulation. (Abelson MB, Udell IJ: Ocular Allergy Update, p 8. Princeton, Excerpta Medica, 1981) |
The inflammatory process is regulated internally by a negative feedback system. The interaction of histamine with mast cell surface histamine receptors elevates cyclic adenosine monophosphate (cAMP) concentrations, thereby “turning off” the mast cell.2 The second messengers, cAMP and cyclic guanosine monophosphate (cGMP), further control mediator release from mast cells and basophils.2 Increasing levels of cAMP block mediator release; increasing levels of cGMP stimulate mediator release. Beta-adrenergic receptor activation enhances cAMP levels; alpha-adrenergic receptor activation diminishes cAMP levels. Prostaglandins act by way of adenyl cyclase to increase cAMP levels. Phosphodiesterase degrades cAMP; thus, phosphodiesterase inhibitors can increase cAMP levels. Cholinergic stimulation results in increasing levels of cGMP and mediator release. Thus, allergic symptoms can be treated by increasing cAMP levels or decreasing cGMP levels. Pharmacologic modulation of these feedback mechanisms may provide a novel method for the treatment of allergic diseases.
PREFORMED MEDIATORS
HISTAMINE
The role of histamine as a mediator of acute inflammation has been clearly established—indeed, histamine represents the prototype of the inflammatory mediators. Histamine is an endogenous substance, widely distributed in mammalian tissue.3 It is stored in the secretory granules of tissue mast cells located primarily in connective tissue associated with blood vessels.4 Histamine is also found in platelets5 and in basophils.6,7 Histamine release can be triggered by both immunologic and nonimmunologic means. Re-exposure of sensitized individuals to an inciting antigen activates cell-bound IgE dimers, inducing mast cell and basophil degranulation and resulting in histamine release. Nonimmunologic mediator release can be induced by agents such as compound 48/80, dextran, and anaphylatoxins (e.g., C3a and C5a), or by trauma.8 Studies in a variety of immediate hypersensitivity models have shown that the release of immediate hypersensitivity mediators is both selective and noncytolytic.
Topical application of compound 48/80 produces the signs and symptoms of ocular allergy (itching, vasodilation, chemosis, and mucous discharge) in the eyes of guinea pigs, rabbits, and humans, and serves as a useful tool in screening anti-inflammatory agents.9 The degranulation of human conjunctival mast cells in response to topically applied compound 48/80, as demonstrated by light microscopy, has been confirmed by transmission electron microscopy.10 Mast cell granules enlarge in response to stimulation by compound 48/80; the number of enlarged granules corresponds to the extent of degranulation. Only cells with pink granules (by alkaline Giemsa stain) and granules enlarged to more than 1 μm showed definite evidence of exocytosis. Caulfield and colleagues, using an in vitro human anti-IgE model, have shown that granules enlarge, coalesce, become amorphous, and then discharge within 1 minute of stimulation.11 Only amorphous granules are capable of discharging their contents.11
Dale and Laidlaw12 used animal models to establish that injection of histamine results in collapsed arterial pressure, hypovolemia, and decreased body temperature. These symptoms are characteristic of traumatic or anaphylactic shock. Local application of histamine to skin results in redness, swelling, and flare from the local axon reflex,13 as well as itching from the stimulation of cutaneous nerve endings.14
To date, three different histamine receptor subtypes have been identified: H1, H2, and H3. Both H1 and H2 receptors have been identified in ocular tissue. When injected intradermally, histamine causes a localized triple response. First is the development of erythema immediately surrounding the injection site. Erythema results from vasodilation mediated by both H1 and H2 receptors.15,16 Second to develop is the cutaneous flare that occurs as an indirect response to stimulation of histamine receptors on afferent nonmyelinated nerve endings. Antidromic nerve conduction (the conduction of impulses in a direction opposite to the normal direction) initiates a reflex arc resulting in the release of various neuropeptides, including substance P and calcitonin gene-related peptide. These neuropeptides have a direct effect on arteriolar vasodilation.17 Finally, wheal results from the exudation of plasma through gaps between the vascular endothelium of postcapillary venules. This is mediated by H1 receptors.18 In addition to the triple response, intradermal injection of histamine causes the sensory response of itching.19
Topical application of histamine to the rabbit or human eye produces the signs and symptoms of an allergic reaction (itching, redness, and swelling) in a dose-dependent fashion.20 H1 receptor activation results in itching21 and redness,20 whereas H2 receptor activation leads to redness.22 The presence of a dual receptor system similar to that of skin has been demonstrated in the eye (Fig. 2). The highly selective H2 receptor agonist dimethylaminopropylisothiourea (dimaprit) produces diffuse vasodilation that can be blocked by cimetidine, an H2 antagonist, but not by antazoline, an H1 antagonist.22 Stimulation of the H1 receptor with the H1 agonist 2-(2-amino-ethyl)thiozole dihydrochloride produces itch and minimal vasodilation, which can be prevented by pheniramine maleate, an H1 antagonist, but not by cimetidine.21
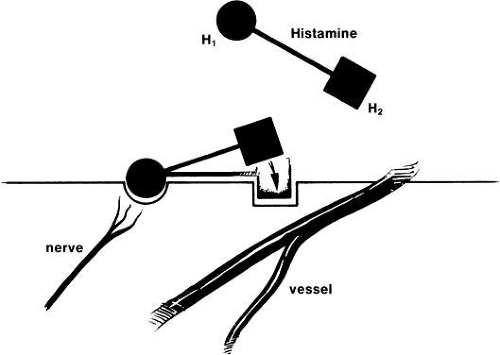 Fig. 2. There are two histamine receptor sites important in allergic conjunctivitis: H1 (itch, burn, plus minimal vasodilation) and H2 (vasodilation). (Abelson MB, Udell IJ: Ocular Allergy Update, p 8. Princeton, Excerpta Medica, 1981) |
Histamine levels have been described in normal human tears (5 to 10 ng/ml). In the past, these levels were not found to be consistently elevated in patients with allergic conjunctivitis, but only in the tears of patients with active vernal keratoconjunctivitis (VKC; 16 ng/ml).23,24 This elevation is probably the result of the extensive mast cell degranulation demonstrated in patients with VKC by light and electron microscopy.25 In addition, patients with VKC have four times as many mast cells in their conjunctiva as normal individuals,25 and the location of these mast cells is more superficial than in the normal conjunctiva.26 Such patients are, therefore, at greater risk for antigenic attack.
A 1990 study examined the presence of histaminase activity in human tears after in vivo conjunctival antigen challenge.27 Inactivation of histaminase resulted in a 15-fold elevation of histamine recovery. Enzyme-inactivated samples had histamine levels of 107.26 ng/ml, whereas untreated samples had histamine levels of 7.07 ng/ml. These results demonstrate the presence of histaminase activity in human tears and suggest that histaminase activity may have confounded the identification of histamine in ocular allergic disorders other than VKC. A 1995 study showed that the degradation of histamine was significantly lower in patients with VKC than controls in both tears and plasma.28
TRYPTASE
Tryptase is a preformed, tetrameric, serine endoprotease found in mast cells. It is stored in abundant quantities in its fully active form. Because tryptase is unique to mast cells, it is an excellent marker for them.29,30 Elevated levels of tryptase have been found in tears of patients after eye-rubbing. In addition, allergen challenge or provocation with compound 48/8031,32 also results in increased levels of tryptase in tears. Tryptase levels appear elevated during the early-phase reaction, but not during the late-phase reaction, after ocular allergen challenge.33 Tryptase is found elevated in patients with VKC, even during the remission phase.32,34
In addition to the eye, tryptase has been found in other biologic fluids such as serum,35 bronchial lavage fluid,36 nasal lavage secretions,37 skin,38 synovial fluid,39 and blisters.40 Tryptase has the ability to potentiate the effect of histamine, activate eosinophils and mast cells, and attract eosinophils and neutrophils. Because of these actions, tryptase inhibitors are being evaluated as potential therapies for asthma and have been shown to have effective anti-inflammatory properties. Mast cell stabilizers have been shown to reduce tryptase levels after allergen challenge.33 Tryptase also has the ability to degrade neuropeptides such as vasoactive intestinal peptide (VIP), peptide histidine-methionine, and calcitonin gene-related peptide. VIP is a prominent bronchodilator, and it is postulated that the destruction of VIP by tryptase results in the increased bronchomotor tone and bronchial hyperresponsiveness41 of asthma. The exact role of tryptase in ocular inflammation is not yet fully understood.
CHYMASE
Two mast cell subtypes have been described: T mast cells, which contain tryptase, and TC mast cells, which contain tryptase and chymase.29 Chymase is a serine endoprotease that is stored, preformed and fully active, in the TC mast cells. Unlike tryptase, chymase is inhibited by plasma proteinase inhibitors.42 The presence of chymase has not yet been demonstrated in the eye, although its presence is suggested by the large number of conjunctival mast cells of the MCtc phenotype.
HEPARIN
Heparin forms complexes with proteases and is released on degranulation of mast cells. Heparin has been shown to have anti-inflammatory properties43 and has been used for the treatment of allergic contact dermatitis,44 but there are only preliminary reports on the use of heparin in ocular allergy.45 Heparin may serve as an endogenous mediator that provides negative feedback on the release of proinflammatory mediators from the mast cell.
EOSINOPHIL CHEMOTACTIC FACTOR OF ANAPHYLAXIS
Eosinophil chemotactic factor of anaphylaxis (ECFA), a preformed mediator of immediate hypersensitivity, was first found in diffusates of guinea pig lung46 and human lung.47 It was subsequently extracted from rat mast cells,48 human leukemic basophils,7 human mast cell-rich lung,48 and nasal polyps.49 The activity of ECFA resides in two closely related tetrapeptides.50
ECFA is released during repeated mast cell challenge and degranulation in severe ocular allergic disease. ECFA is a potent chemotactic factor for eosinophils; therefore, the release of large amounts of ECFA is responsible for conjunctival eosinophilia.51 The presence of eosinophils in a conjunctival scraping is always indicative of ocular allergy. In fact, a conjunctival scraping containing two or more eosinophils per high-power field is considered diagnostic of ocular allergy.51 However, by definition, eosinophils do not have to be present in conjunctival scrapings to make the clinical diagnosis of ocular allergic disease, because they may be located only deep in conjunctival tissues.
ECFA is more selective than complementderived fragments,52 cocytotoxin,53 and peptides extracted from neoplasms.54 Interaction with ECFA reduces the chemotactic responsiveness of eosinophils,55 thereby retaining these cells at a specific site for regulatory purposes. Eosinophils release a number of substances that modulate the mast cell response and limit mediator activity: histaminase inactivates histamine,56,57,58 phospholipase inactivates platelet-activating factor,59 and aryl sulfatase inactivates leukotrienes.60,61,62
EOSINOPHIL GRANULE MAJOR BASIC PROTEIN
In addition to their beneficial role as modulators of inflammation, eosinophils are also responsible for tissue destruction, as noted in the cytotoxic effects of certain granular proteins on parasites and mammalian tissues.63 Eosinophil granule major basic protein (EMBP) accounts for more than 50% of the eosinophil granule protein and 25% of the total cellular protein.64,65
EMBP is a strongly cationic molecule with a molecular weight of 9,300 in humans and 11,000 in guinea pigs.66,67 A harmful role has been suggested for the eosinophil because of the markedly increased levels of EMBP is asthmatic sputum,68 and the ability of EMBP to mimic the pathologic changes seen in asthma.69 It has been demonstrated to elicit mast cell70 and basophil71 degranulation. Both VKC and contact lens-associated giant papillary conjunctivitis are associated with marked mast cell degranulation and eosinophil infiltration,72,73 providing further evidence that eosinophils play a role in tissue damage.
Increased tear levels of both EMBP and Charcot-Leyden crystal protein have been detected in patients with VKC. The increased EMBP levels appear to correspond to the severity of the disease.74 Also, immunofluorescence of conjunctival tissue from normal patients revealed little or no EMBP, whereas specimens from patients with VKC and contact lens-associated giant papillary conjunctivitis revealed significant EMBP deposition.75 However, in this study no correlation between the intensity of EMBP deposition and the severity of disease was documented. Thus, it appears that both VKC and contact lens-associated giant papillary conjunctivitis are characterized by eosinophil degranulation with the release of EMBP and other cytotoxic granule proteins that may further stimulate mast cell degranulation.75
The release of EMBP, a powerful epithelial toxic compound, may account for keratitis and shield ulcers in VKC. EMBP deposits were identified by immunofluorescence in the base and the mucus plug in two corneal shield ulcers removed by superficial keratotomy from two patients with VKC. EMBP may also contribute to sustained mast cell degranulation and thus the severe and protracted process associated with this condition.76
PLATELET-ACTIVATING FACTOR
Benveniste and colleagues named and characterized platelet-activating factor (PAF) as a phospholipase A2 (PLA2)-sensitive phospholipid,77,78,79,80 identified as 1-alkyl-2(R)-acetyl-glycero-3-phosphorylcholine.80,81 PAF is the most potent eosinophil chemotactic factor known and is approximately 100 times more effective than ECFA or leukotriene B4 (LTB4). It has been reported to mediate inflammation and vascular permeability.82 PAF has also been implicated in other pathophysiologic processes,83 such as aggregating platelets, chemotaxis and degradation of eosinophils and neutrophils, bronchoconstriction, increased bronchial responsiveness, and hypotension. Rabbit basophils have been shown to release PAF by an IgE-dependent process.84 Mast cells, eosinophils, monocytes, basophils, polymorphonuclear leukocytes, and macrophages have also been demonstrated to release PAF when stimulated with IgE, a calcium ionophore, or zymosan particles.84
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


