Innervation of the Extraocular Muscles
Michael X. Repka
The third, fourth, and sixth cranial nerves innervate six extraocular muscles in each orbit. These muscles are responsible for controlling the ocular rotations in horizontal, vertical, and torsional directions. The third cranial nerve also provides parasympathetic motor innervation to the intrinsic eye muscles, the ciliary muscle, and the iris sphincter muscle. Although these nerves originate at separate locations in the brainstem, they converge in the subarachnoid space and become closely associated in the cavernous sinus, superior orbital fissure, and posterior orbit (Fig. 3.1). From the cavernous sinus to the eye, they may be simultaneously involved in pathological processes.
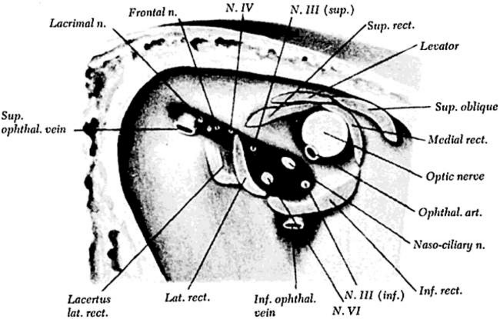 Figure 3.1 Superior orbital fissure and annulus of Zinn. Cranial nerves III and VI enter the orbit within the muscle cone, but cranial nerve IV enters lateral to the annulus of Zinn. Source: Duke-Elder S: System of Ophthalmology, Vol 2: The Anatomy of the Visual System. London: Kimpton, 1961:420 |
Oculomotor Nerve
Nucleus
The oculomotor nerve is the largest of the ocular motor nerves. It contains some 15,000 axons, including motor fibers and parasympathetic motor fibers. The nucleus of the third cranial nerve is located in the inferior periaqueductal gray matter of the mesencephalon. It is a complex of paired and midline motor cells. These cells are found from the posterior commissure superiorly to the pontomesencephalic junction caudally (Fig. 3.2) in the dorsal midbrain. Classically, this complex has been described with four paired subnuclei supplying innervation to the inferior, medial, and superior rectus muscles and to the inferior oblique muscle. There is a single caudal, dorsal midline nucleus providing innervation to the levator muscle. Innervation of the extraocular muscle from the paired nuclei is ipsilateral for the inferior and medial rectus muscles and the inferior oblique muscle. However, innervation of the superior rectus is contralateral. The motor fibers to the superior rectus decussate within the third nerve nucleus and join the fascicle of the contralateral oculomotor nerve.
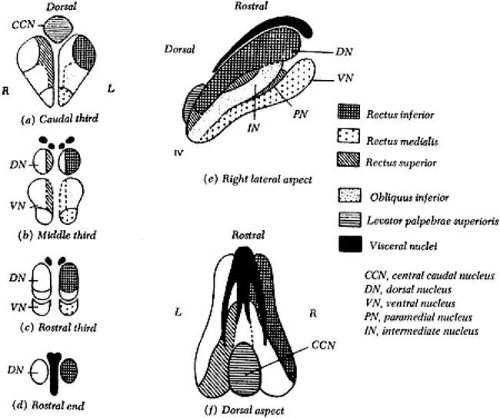 Figure 3.2 Warwick’s arrangement of the III cranial nerve subnuclei, illustrating the pattern of motor cells in the brainstem that send fibers to each extraocular muscle in the oculomotor nuclei of the monkey. The medial rectus subnucleus has been divided into three regions. The cells of the nucleus identified by Warwick represent those called “A” motor neurons by Büttner-Ennever (5). Source: Warwick R: Representation of the extraocular muscles in the oculomotor nuclei of the monkey. J Comp Neurol 1953;98:449–503 |
The organizational scheme of the oculomotor nucleus was proposed by Warwick.1 This hypothesis was substantiated by using retrograde degeneration techniques in monkeys. Substantiation of Warwick’s hypothesis in humans relies on autopsy findings and neuroimaging results.2,3,4 Recent research in monkeys has suggested that the medial rectus muscle may not have only one subnucleus, as suggested by Warwick, but may receive innervation from three separate regions within the oculomotor nerve complex, which have been labeled A, B, and C.5
In addition to the extraocular muscle motor fibers, the Edinger-Westphal nuclei supply the parasympathetic preganglionic neurons that project to the ciliary ganglion. These nuclei are positioned dorsal and medial to the muscle motor nuclei. They are composed of two cell groups: a medial unpaired column and a lateral paired column. All evidence suggests that these fibers project ipsilaterally. There is a second source of parasympathetic fibers projecting to the ciliary ganglion in monkeys. Burde and Loewy6 showed the source of these fibers to be Perlia’s nucleus. This nucleus is found in the middle third of the oculomotor complex, positioned dorsally. The role of this structure in humans is unknown.
Fascicle
The fascicular portion of the oculomotor nerve descends as its moves ventrally and laterally through the mesencephalon (Fig. 3.3). The fascicles pass through the medial longitudinal fasciculus, the red nucleus, and the medial portion of the cerebral peduncle. During the passage through the ventral midbrain, there appears to be topographic organization of the fascicles so that a divisional paresis is possible, even with intrinsic lesions.4 The oculomotor nerve exits the brainstem into the interpeduncular fossa as a horizontal arrangement of multiple fiber bundles that rapidly coalesce to form the subarachnoid portion of the third cranial nerve.
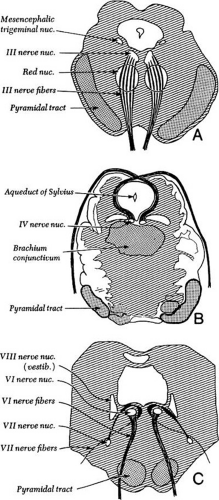 Figure 3.3 Cross-section of the brainstem through superior colliculus, showing cranial nerve III nuclei with intramedullary fibers. |
Nerve
The oculomotor nerve exits the brainstem coursing downward and laterally through the intracranial, prepontine space, reaching the top of the clivus at a point just lateral to the posterior clinoid process. During the passage through the subarachnoid space, the nerve passes just below the posterior cerebral artery and above the superior cerebellar artery (Fig. 3.4). After passing between these arteries, the nerve travels adjacent to the posterior communicating artery. The close apposition of the third nerve to these vascular structures accounts for the frequent involvement of this nerve by vascular anomalies, such as an aneurysm of the posterior communicating artery.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


