Dietary Prevention and Treatment of Ocular Disease
Dietary Prevention and Treatment of Ocular Disease
Billy R. Hammond Jr.
Eric Lien
INTRODUCTION
Training in nutrition in most allopathic medical schools is minimal1 and limited to a focus on diet as a means of preventing, largely, metabolic diseases. The diet as drug (“a pill for every ill”) approach likely originated with the study of scurvy, which was inevitably fatal if not treated. Scurvy, for instance, afflicted sailors (who had a limited range of food supplies) to the extent that it often threatened the success of their missions. Treatment of scurvy by diet is nearly as old as recorded history but it was not recognized until the 15 to 16th century that a specific ingredient within food was responsible for its curative value (see the review by Carpenter2). This finding inspired generations to conclude that isolated elements of food could prevent, or sometimes cure, disease. After the success of vitamin C, other nutrients were found to prevent and also seemingly “cure” serious metabolic diseases: iodine and goiter, vitamin D and rickets, thiamine and beri beri, protein and kwashiorkor, vitamin A and night blindness, etc. The single-agent approach was encouraged by the advent of the pharmaceutical industry, in which one therapeutic agent successfully treats specific symptoms. For example, acetylsalicylic acid could reduce fever and relieve pain from arthritis, penicillin could cure numerous bacterial infections, and insulin could treat diabetes.
Unlike pharmaceuticals, however, using single nutrients as a treatment for most common ailments is nearly always limited. Complex mechanisms support all cellular functions and require the participation of numerous dietary components or their derivatives. The successful functioning of any organ system requires the highly integrated action of a variety of nutrients. Successful study of these interactions, however, is often challenging when applied to degenerative diseases (discussed later in this chapter) that are slow in onset and require years of suboptimal intake to become manifest. Cross-sectional studies tend to evaluate dietary intake of a large number of individuals and correlate specific dietary intake to disease. Such data represent only a “snap shot” in time. Dietary habits may change over a period of time, and thus beneficial or detrimental dietary-component intake may not be stable. Prospective studies provide firmer evidence of cause and effect. Tracking dietary intake over an extended period of time, however, is also exceptionally difficult. Some studies recruit individuals prospectively, follow dietary intake and disease incidence for a predetermined time and evaluate the effect of specific dietary components on the development of disease. Although these data are stronger than cross-sectional data, they lack the strength of administration of specific components (or placebo) to a well-matched group of individuals. The randomized placebo-controlled trial provides the most convincing evidence of cause and effect. These studies, however, are extremely complex and costly and therefore require significant funding. This is often accomplished by companies that are interested in promoting the ingredient they produce. These type of commercial interests have, over the last decade or so, prompted many academic journals to be more careful in the full disclosure of financial conflicts.
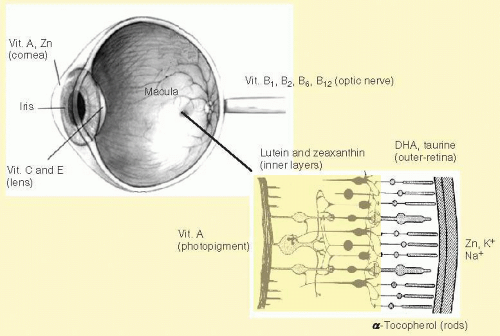
Commercial input can also be reflected simply in the availability of nutrients that can be used for study. For example, the Age-Related Eye Disease Study (AREDS) was the first large-scale study on nutrition that was ever conducted by the National Eye Institute.3 The eye is, of course, replete with nutrients and other dietary components (see Fig. 34.1), and their interactions are complex. AREDS started enrolling subjects (55 to 80 years) at 11 different sites across the United States in 1992. This recruitment continued until 1998 resulting in 3,557 participants that were studied (vision exams every 6 months) with an average follow-up of 6.3 years. The aim of the study was to assess whether an intervention with vitamins/zinc could alter progression of late-stage retinal and lens disease. The cocktail of nutrients that was used included beta-carotene as a major portion of the formulation. There are approximately 20 dietary carotenoids, including beta-carotene, that are found in the serum and body. Only lutein and zeaxanthin (and an intermediary isomer, meso-zeaxanthin), however, are normally found in the retina. Beta-carotene (not found in the retina under normal circumstances but a recognized antioxidant) was included not because of its presence in the eye, but rather because L and Z were not commercially available (most of the carotenoids in the serum are not) (see Hammond and Johnson4). AREDs II is now including both lutein and zeaxanthin and omega-3 fatty acids in the formulation.5
FIG. 34.1 The eye is a nutritionally responsive tissue: some examples of the many nutrients found throughout the eye.
Although literally thousands of studies have examined the role of diet and degenerative disease, the results can at best be described as mixed. There are likely several reasons for this. The first reason is that most research, like the earlier research on deficiency disease, follows the “single nutrient” model. Food, like disease itself, represents a complex of factors, and attempting to disentangle one compound from such a complex mix is extremely challenging (see the recent review by Mares et al.6). One can, of course, do interventions: supplement a nutrient and look at the effects on disease incidence and/or progression. Invariably, however, such interventions must be done very late (often after decades of irreversible damage) and are inconsistent with the idea that nutrients prevent damage over a lifespan.* They must be conducted in elderly individuals because it is necessary for a small group of the patients to actually get the disease in order to link an etiological factor to its development. Cross-sectional studies can be done retrospectively, but assessing diet far in the past is also very challenging.
These reasons, among many others, are likely why epidemiology, as a discipline, has not been particularly successful at identifying clear risk and protective factors for most degenerative diseases. The most success has come from the study of variables that are easily quantified and tend to be temporally stable (like smoking tobacco). In order then to accurately evaluate the evidence for diet as a means of preventing deficiency or degenerative diseases of the eye, or as a palliative, evidence from a wide variety of sources must be considered.
PREVENTION OF DEFICIENCY DISEASES OF THE VISUAL SYSTEM (ESSENTIAL AND CONDITIONALLY ESSENTIAL NUTRIENTS)
Essential Nutrients
The concept of the essentiality of specific food components developed dramatically during the first half of the 20th century, and by 1950 scientists defined five criteria for essentiality9,10:
1. The dietary component is required for growth, health, and survival.
2. Characteristic signs of deficiency disease (and ultimately death) result from inadequate intake of the nutrient.
3. Growth retardation and specific signs of deficiency are alleviated in a dose-related manner in response to increasing dietary levels of the nutrient.
4. The substance is not endogenously synthesized.
5. Signs of deficiency and growth failure are prevented only by the nutrient or its precursors and not by other substances.
Typically nutritional deficiencies evolve through a specific sequence. During early stages of deprivation, blood and tissue levels are maintained by use of reserves from specific storage organs (e.g., hepatic vitamin A during periods of reduced vitamin A intake). As nutrient stores are depleted, biochemical alterations occur, although clinical symptoms may not be apparent. Finally, clinical symptoms, as well as altered biochemical pathways, become manifest with the development of a full-blown nutritional deficiency syndrome.11 The above criteria serve to define essential nutrients when a deficiency state becomes obvious in a relatively short period of time, such as scurvy in the crews of sailing vessels. Overt vitamin C deficiency characterized by lethargy, dermal manifestations, and bleeding from the gums becomes manifest after a few months of very low vitamin C intake, such as during extended ocean voyages. Institution of a modest daily dose of citrus juice (generally lime juice) produces a rapid resolution of deficiency symptoms.
In contrast to the obvious symptoms of a nutritional deficiency as outlined by the five points above, a number of trace elements fail to meet these criteria, and yet are clearly essential. Elimination of dietary sources of specific elements is difficult in experimental animals, and in humans classic signs of deficiency occur only after prolonged periods of dietary deprivation. The doctrine of the first half of the 20th century that deficiency of an essential nutrient must result in severe symptoms, minimally resulting in growth retardation (if not death), was clearly not appropriate for a number of trace elements, and the definition of an essential nutrient was broadened. The essentiality of trace elements (such as zinc, copper, manganese, and selenium) rests on the three criteria of induction, prevention, and resolution: (1) induction of specific symptoms during a deficiency state, (2) prevention of deficiency symptoms when the element is present in the diet at required levels, and (3) resolution of specific deficiency symptoms following introduction of a diet replete with the element under study.11 These deficiencies may be difficult to generate and have complex relationships with the overall diet. For example, zinc is a cofactor for literally hundreds of enzymes, yet its essentiality was recognized only under dietary conditions which limited its bioavailability by consumption of diets high in components that inhibit zinc absorption, such as phytate and calcium.12 Zinc deficiency, characterized by retarded growth and sexual manifestations (e.g., dysmenorrhea), developed in adolescents when they consumed diets containing these zinc-absorption antagonists, although their diets also contained zinc levels that were generally considered to be adequate. Resolution of these deficiency signs required high oral zinc intake.13,14 This and other examples demonstrate that the stringent definition of nutritional deficiency must be complemented by knowledge of the complex interactions between the nutrient under consideration and other dietary components.
Conditionally Essential Nutrients
Humans are capable of de novo synthesis of many of the biochemicals that they require (e.g., nonessential amino acids, neurotransmitters, and intermediary metabolites). However, in certain physiological or pathological states, specific biochemicals may not be synthesized in sufficient quantity for health maintenance and a dietary source must be consumed. Such nutrients are termed “conditionally essential.” Four mechanisms may lead to inadequate endogenous synthesis15:
1. A polymorphism that blocks a biosynthetic pathway.
2. Disease of an organ which leads to loss of enzymatic activity.
3. Developmental immaturity in preterm or term neonates, when a metabolic pathway involves enzymes that mature late in fetal development or early in infancy.
4. Conditions such as total parenteral nutrition (TPN) in which liver function may be impaired/bypassed and the products of hepatic enzymes are not produced at adequate levels.
For example, premature infants are incapable of synthesizing several amino acids at adequate rates for their requirements that are nonessential in mature individuals.16 The requirements for these amino acids must be met by dietary means, and thus these amino acids are conditionally essential for preterm infants. In addition, the synthesis of the long-chain polyunsaturated fatty acids (LCPUFA), docosahexaenoic acid (DHA), and arachidonic acid (ARA) is limited in preterm infants, and these LCPUFAs may be considered conditionally essential.17 Another example involves individuals receiving TPN for extended periods. Choline is synthesized from its precursor, methionine, by the liver, and parenteral provision of methionine will decrease hepatic availability of this amino acid. This type of nutrition may result in low levels of plasma choline and is associated with hepatic steatosis.18
Night Blindness, Xerophthalmia, and Vitamin A
Although rare in developed countries, vitamin A deficiency remains the leading cause of childhood blindness in the world. Some 250,000 children go blind annually from deficiencies in vitamin A, with about half of those dying within the first year.19,20,21 Dietary vitamin A is basic to the eye’s physiology.7 For instance, rhodopsin resides within the disc membranes of rod outer segments and is a complex of 11-cis retinal, the visually active form of vitamin A, covalently bound to the transmembrane protein opsin. 11-Cis retinal is located in an intermembrane section of rhodopsin, and the visual cascade is initiated upon absorption of a photon of light by this conformation of retinal, converting it to all-trans retinal. The isomerization of retinal results in a conformational shift of rhodopsin and all-trans retinal-opsin no longer fits in the compact membrane binding site of 11-cis retinal-opsin. Rhodopsin then passes through a series of unstable intermediates to generate metarhodopsin I and metarhodopsin II.22 These confirmations are in equilibrium, with metarhodopsin II being the metabolically active conformation. The generation of metarhodopsin II results in the activation of transducin, a G-protein family member, which in turn activates cGMP phosphodiesterase. Upon the cGMP phosphodiesterase catalyzed hydrolysis of cGMP, the cGMP-gated sodium channel is closed and the cell becomes hyperpolarized.23 Similar complexes of 11-cis retinal with three different opsin-related proteins exist within cone outer segments. Excitation of the cone cells is wavelength dependent and relates to the specific protein-retinal complex that individual cone cells possess.
The first clinical manifestation of vitamin A deficiency is night blindness, caused by prolonged recovery of rods following excitation.24 This alteration of photoreceptor function is due to 11-cis retinal depletion in the rod outer segment. Vitamin A in its retinoic acid form is essential for ocular activities which are not related to retinal function.25,26 Low retinoic acid status limits goblet cell function in the conjunctiva with the result of a dry corneal surface. Bitot’s spots are formed from dead microbial and epithelial cells, and in more severe vitamin A deficiency this is followed by conjunctival xerosis, characterized by a dry and rough cornea. These progressive stages of vitamin A deficiency are reversible as long as corneal ulcerations have not occurred. However, once ulceration takes place the process is irreversible and vision becomes permanently affected.26
Dietary Sources of Vitamin A
Vitamin A has two dietary sources. One is preformed vitamin A from animal origins and is present in high concentrations in human milk, whole cow’s milk, eggs, and liver. These animal sources of vitamin A are in the chemical form of a fatty acid ester, retinyl palmitate. In the small intestine, this ester is hydrolyzed to release retinol, the alcohol congener of vitamin A.27 Margarine as well as nonfat and low-fat dairy products is often fortified with vitamin A in its vitamin A palmitate form. Highly pigmented vegetables such as carrots, squash, cantaloupe, broccoli, and spinach provide pro-vitamin A carotenoids such as beta-carotene, alpha-carotene, and beta-cryptoxanthin. Carotenes can be converted to active vitamin A metabolites with beta-carotene being the carotenoid most efficiently converted to vitamin A.28 The Institute of Medicine, National Academy of Sciences, states that 12 µg beta-carotene is converted to 1 µg of retinol,29 although more recent data suggest that the conversion factor for both vegetables and fruits is less than this value.30,31Table 34.1 provides a brief summary of the level of preformed vitamin A and of beta-carotene in selected foods.32 Once absorbed, vitamin A, generated from either animal or plant sources, is converted to several lipid-soluble molecules with a variety of activities.29 As discussed above, the derivatives active in the visual system are 11-cis retinal, an aldehyde which is an integral component of rhodopsin, and retinoic acid, essential for epithelial cell function and maintenance of cornea and conjunctival membranes (thus essential for preventing xerophthalmia).19 In addition to its functions within the eye, vitamin A is essential for proper expression of genes that code for structural proteins such as skin keratins, enzymes, and retinol-binding protein.
Vitamin A is the only nutrient that is clearly defined as essential for vision. Dietary sources of this vitamin are required for both retinal function and maintenance of epithelial cells. Dietary requirements for vitamin A are based on the maintenance of adequate stores of vitamin A, and these requirements are sufficient to ensure prevention of night blindness. Table 34.2 provides the Recommended Daily Allowances (RDAs) for ages throughout the life span as set by the Institute of Medicine, National Academy of Sciences.33 The RDAs are the average daily dietary intake level sufficient to meet the nutrient requirements of nearly all (97% to 98%) healthy individuals in a group. If data are insufficient to develop an RDA, the Institute of Medicine will develop a value termed “adequate intake” (AI). For healthy term infants, this value is based on the daily nutrient consumption of breast-fed children. For other age groups, an AI is believed to cover the needs of all healthy individuals in the group, but a lack of data prevents an RDA from being developed.
TABLE 34-1 Dietary Sources of Vitamin A and Beta-Carotene
From Agricultural Research Service, United States Department of Agriculture. USDA National Nutrient Database for Standard Reference. 2011. Available from http://www.ars.usda.gov/main/site_main. htm?modecode=12-35-45-00. Accessed December 12, 2012.
TABLE 34-2 Vitamin A, Recommended Daily Allowances
Modified from Panel on Micronutrients, et al. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc. Washington, DC: National Academies Press; 2001.
PREVENTION OF DEGENERATIVE EYE DISEASE
Nutrients are defined34 as dietary elements that cannot be synthesized de novo and whose absence leads to a deficiency disease (although there are some curious exceptions from this definition such as vitamin E). Many elements of the diet, however, appear to prevent long-term damage and are therefore likely important in preventing age-related deterioration. For example, the retina is likely damaged by exposure to light with wavelengths from about 400 to 500 nm. Such light is not effectively screened by the anterior lenses of the eye yet is sufficiently energetic to initiate photochemical damage. There is empirical evidence that lutein (L) and zeaxanthin (Z) within the inner layers of the retina (i.e., macular pigment, MP) can screen such light and prevent the resultant actinic damage.35 Subjects vary dramatically36 in the amount of MP they possess ranging from near zero (100% transmission) to over one log unit at peak absorbance (<10% transmission). Such a dramatic difference, when considered over a lifespan, can significantly change the deterioration rate of the tissue (analogous to how melanin in darker skin protects from light damage). Since pathological changes to neural tissue tend to cascade,37 increased protection even late in life could yield significant benefit. Food components such as L and Z are not defined as essential nutrients, but have beneficial activities. These components have received increasing interest and scrutiny, with authoritative bodies recognizing that the traditional definitions for nutrients that maintain optimal health should be broadened.38,39,40 The field of nutrition traditionally concentrated on the prevention of specific nutritional deficiencies but has now moved beyond this narrow focus in order to address food component-mediated reduction of disease risk. A distinction exists between absolutely essential nutrients and beneficial food components that reduce the risk of chronic diseases, yet both contribute to optimal health.41
Many food components have been studied as visual prophylaxes, and a complete review would require a lengthy text. Hence, in this review, our focus will be on three: fatty acids, antioxidants, and taurine.
FIG. 34.2 Biosynthesis of lc-PUFAs.
Fatty Acids
Fatty Acid Classes
Fatty acids are carboxylic acids with a linear tail of carbon atoms. These linear chains may be of varying degrees of unsaturation containing no double bonds (saturated), one double bond (monounsaturated), or multiple double bonds (polyunsaturated). In addition to double bonds, fatty acids are characterized by the number of carbons contained in the linear chain from the carboxyl group (alpha position) to the methyl end (omega position). Thus, C16:0 indicates that the fatty acid is 16 carbons long and has no double bonds (palmitic acid). The first point of unsaturation is designated by the number of carbons from the methyl end of the fatty acid chain. Oleic acid is a monounsaturated fatty acid with its single point of unsaturation nine carbons from the methyl end and is designated C18:1 n-9 (or C18:1ω9). Humans are capable of synthesizing saturated fatty acids and monounsaturated fatty acids (with the point of unsaturation at the n-9 position). However, they cannot synthesize polyunsaturated fatty acids of the n-3 and n-6 families. These types of fatty acids serve critical functions within the body and are essential dietary components.
Linoleic acid (LA, C18:2 n-6) is the parent fatty acid of the n-6 family whereas alpha-linolenic acid (ALA, C18:3 n-3) is the parent of the n-3 family. These fatty acids are converted by a series of enzymatic reactions to long-chain polyunsaturated fatty acids (polyunsaturated fatty acids which are 20 or 22 carbons in length). For example, LA is converted to arachidonic acid (ARA, C20:4 n-6), the precursor to numerous prostaglandins, leukotrienes, and other highly bioactive mediators. In an analogous manner, ALA is converted to eicosapentaenoic acid (EPA, C20:5 n-3). Although the structure of EPA is very similar to ARA (EPA contains one additional double bond in the n-3 position), the biological activities of EPA are strikingly different from ARA.42 As seen in Figure 34.2, LA and ALA compete for the same chain elongation and desaturation enzymes, and the production rates of ARA and EPA are very dependent on substrate concentration. Because humans cannot insert double bonds at either the n-3 or n-6 position, these two families of fatty acids are both nutritionally essential.43
Further chain elongation and desaturation of EPA results in the synthesis of DHA, derived by a circuitous route as outlined in Figure 34.2. Delta 6 desaturase is an enzyme involved in the synthesis of ARA from LA and the synthesis of EPA from ALA. In addition, the same desaturase is required for the synthesis of DHA from EPA. The competition for this common enzyme, as well as the relatively low concentrations of EPA compared with the C18 substrates LA and ALA, result in only marginal production of DHA from its precursors. Most published studies in adult humans report that an increase in dietary ALA results in increased plasma and RBC EPA levels.44 In contrast, DHA status is not increased by a greater consumption of ALA, demonstrating that increased DHA status must occur predominantly by consumption of preformed DHA.44 Stable isotope studies in adults have demonstrated that the conversion of ALA to DHA is strikingly inefficient. Pawlosky et al.45 found that only 0.2% of orally consumed ALA is converted to EPA and 0.05% is converted to DHA.
The fatty acid status of term infants is a useful example of the complex interactions of the C18 essential fatty acids, LA and ALA, and their LCPUFA derivatives. Manipulation of infant formula LA to ALA ratios demonstrates that only marginal increases in DHA occur when term infants consume relatively high levels of its precursor, ALA.46,47 Due to the competition of ALA and LA for the same chain elongation and desaturation enzymes, high levels of dietary ALA suppress conversion of LA to ARA, resulting in reduced plasma ARA levels (essential for growth of infants) and growth rates.46 Human milk contains preformed DHA,48 and breast-fed infants have much higher plasma and RBC DHA levels than do infants fed any combination of ALA and LA in infant formula.47,49 Thus, the appropriate means of supporting DHA levels in infants is through dietary consumption of DHA itself, not of its precursors. Inefficient conversion of ALA to DHA is apparent in both infants and adults, and therefore dietary sources of DHA are necessary at all ages if maintaining high levels of circulating DHA is an appropriate goal.
Retinal DHA
DHA is the predominant fatty acid of rod and cone outer segments, where it constitutes up to 50% of total fatty acids.50,51 The retina has mechanisms to specifically sequester DHA from the plasma, insert it into the photoreceptor outer segment membrane, and then recycle it once the disc is lost from the photoreceptor.52 The retinal pigment epithelium (RPE) plays a critical role in the replenishment of shed discs of rods or disc-like elements of cones. As DHA-rich phospholipid membranes are shed, they are phagocytized by the RPE and the DHA is returned to the photoreceptor for regeneration of outer segment discs.52,53 This efficient short-loop recycling process is critical to the maintenance of retinal function due to the rapid loss and resynthesis of rod discs.54
Rod and cone outer segment DHA is essential for the efficient operation of the visual cascade. DHA-rich membranes are highly unstructured and very disordered,55 while membranes containing shorter and more saturated fatty acids are of a more uniform structure. Each point of unsaturation in a fatty acid results in the introduction of a 37-degree angle in the fatty acid chain.56 Diets low in DHA and high in n-6 fatty acids result in replacement of DHA with less unsaturated fatty acids, such as docosapentaenoic acid n-6 (DPA n-6, C22:5 n-6)57 with the result of altered membrane fluidity.58 Optimal function of rhodopsin is sensitive to such changes in membrane composition.58
The location of rhodopsin’s 11-cis retinal is within the disc outer segment phospholipid membrane, and therefore the initiation of the visual signal occurs within this membrane (cone outer segments provide a similar placement for 11-cis retinal and the opsin-like proteins of cone visual pigments). A photon of light converts rhodopsin to metarhodopsin I and metarhodopsin II, which are in equilibrium with each other. Metarhodopsin II is the active form, and the equilibrium is driven to this product in DHA-rich membranes. Metarhodopsin II formation is progressively reduced in membranes containing more saturated and shorter chain fatty acids.59,60 Metarhodopsin II subsequently binds to transducin (a hetero-trimeric G protein). This is a very rapid step, due to the high concentration of transducin in the membrane and also the highly fluid nature of the DHA-rich membrane. Once the transducin-metarhodopsin II complex is formed, two of the transducin subunits are lost and the active transducin binds to cGMP phosphodiesterase. The rate of formation of the metarhodopsin II/transducin complex61 and affinity of metarhodopsin II for transducin62 are related to the membrane environment, with high affinity in DHA-containing membranes and lower affinity in more saturated membranes. Activated cGMP phosphodiesterase hydrolyzes cGMP to 5’GMP with the resultant closing of the outer segment cGMP-gated sodium channel.63 In an in vitro model system, the phosphodiesterase activity was 50% lower in membranes containing DPA n-6 compared with membranes reconstituted with DHA.60
The above steps constitute the initiation of the visual signal. Niu et al.64 evaluated the initial steps of the visual cascade in rats receiving n-3 sufficient or deficient diets for two generations. The deficient animals had 80% less rod outer segment DHA compared with n-3 replete animals, with replacement by DPA n-6. The replacement of DHA by DPA n-6 resulted in the diminution of visual signaling as demonstrated by slower metarhodopsin II formation, reduced rhodopsin-transducin coupling and reduced cGMP phosphodiesterase activity. These results are in excellent agreement with in vivo retinal electroretinogram (ERG) results from animals receiving diets free of n-3 fatty acids compared with n-3 fatty acid-replete animals. ERGs from animals deficient in n-3 fatty acid have reduced signal amplitude and a lag time in signal development compared with n-3 fatty acid-replete controls, as would be predicted from inefficiencies in the initial steps of the visual cascade.59,65
Developmental Role of DHA
DHA-rich membranes support optimal signal transduction from formation of metarhodopsin II to cell polarization (closing of the cGMP sodium channel of the rod and cone outer segments). Differences in DHA intake and status may play a role in visual development during the neonatal period. Human milk contains DHA, with the concentration being dependent on maternal diet. For example, a 6-fold difference in milk DHA concentration exists between women consuming diets relatively low in fish (e.g., internal locations in the US and Canada) versus women consuming diets rich in fish (e.g., Tokyo, Japan).48 In addition to the presence of DHA in human milk, many but not all infant formulas are supplemented with this LCPUFA. Studies in infants have evaluated the effects of DHA status on visual development. For example, in a cross-sectional study, Innis et al.66 found a positive correlation between infant RBC DHA concentration at 2 months of age and visual acuity at both 2 and 12 months (but not at 4 and 6 months of age). Makrides et al.67 compared visual evoked potential (VEP) and RBC DHA profiles in breast-fed and formula-fed infants at approximately 4 months of age (formulas did not contain LCPUFAs); VEP was more mature in the breast-fed group than in the formula group, and VEP was correlated with RBC DHA status. Lauritzen et al.68 evaluated visual acuity in infants whose lactating mothers received a fish oil supplement or a placebo control. Although there was no difference between groups, the infants’ visual acuity was positively correlated to their RBC DHA status at 4 months of age. These results suggest a relationship between visual development and dietary DHA, but do not conclusively prove cause and effect.
The results of clinical studies have provided a rationale for the fortification of formulas for both preterm and term infants. Initial studies in preterm infants employed formulas supplemented with fish oil containing more EPA than DHA.69,70 This is a departure from the human milk model, which is generally dominant in DHA.48,71 Results from these initial trials indicated that infants receiving formulas enriched in n-3 fatty acids had more mature vision (measured by acuity card preferential looking test or ERG) than infants in the control groups. Subsequently, formula manufacturers developed formulas that are fortified with DHA derived predominantly from fermentation sources or high-DHA fish oil with minimal or no EPA. Many of these formulas also contain ARA.
Some, but not all, trials in preterm infants have demonstrated superior vision in infants receiving formulas supplemented with LCPUFAs compared with controls. An initial meta-analysis72 reported a beneficial effect of supplemented formulas on visual development, although a more recent Cochrane review did not report a consistent positive effect on visual development.73 One consideration for preterm infants relates to the elevated requirement for DHA during the first few months of life. Since the infant is deprived of the intrauterine environment, requirements for DHA may be higher than for term infants. Smithers et al.74 compared formulas containing a typical amount of DHA (0.35%)* versus a higher level (approximately 1.0%). At 2 months corrected age, the VEP of the two groups did not differ, while at 4 months, the VEP was superior in the high-DHA group than in the low DHA.
Numerous studies have evaluated visual development of term infants receiving formulas supplemented with DHA or control formulas. The formulas utilized in these studies varied substantially in the DHA levels, proportions of ALA and LA (precursors of DHA and ARA), and presence or absence of preformed ARA. Some studies have employed formulas exceptionally low in DHA (<0.2%), and it is not surprising that the results from such studies are inconclusive.75 A Cochrane review did not find a consistent improvement in visual acuity in term infants receiving formulas supplemented with DHA with or without ARA76; however, due to the many differences cited above, direct comparison of the available study results is difficult. In order to limit these variables and establish an efficacious DHA addition level, Birch et al.77 evaluated four similar formulas, differing only in their DHA levels (0%, 0.32%, 0.64%, and 0.96%; all formula supplemented with DHA also contained 0.64% ARA). VEP was determined at 1.5, 4, 9, and 12 months of age. The group receiving the unsupplemented formula had significantly poorer VEP acuity at all ages than any of the groups receiving DHA-containing formulas. The three supplemented groups had similar VEP acuity, indicating that a DHA supplementation level of 0.32% in term infants is sufficient to support visual development. Importantly, this DHA level is similar to the global mean of human milk DHA.58
Supply of DHA to young children is also important at time periods beyond the first few months of life. After 6 months of age, plasma and RBC DHA levels in breast-fed infants typically decrease due to decreased consumption of breast milk as a proportion of total diet (weaning foods are generally relatively poor sources of DHA). Hoffman et al.78 evaluated the (VEP) response of breast-fed infants receiving complementary foods supplemented with DHA-enriched egg yolk or similar foods without egg yolk at 1 year of age. DHA status (RBC DHA) increased in the children receiving DHA-supplemented complementary foods, whereas it decreased in the control group. VEP acuity was superior in the supplemented group compared with the control group, and VEP acuity was correlated with RBC DHA. Support of DHA levels during the second 6 months of life may also be achieved by weaning breast-fed infants to a DHA-enriched formula. As with infants receiving DHA-enriched weaning foods, infants weaned to a DHA-containing formula had both higher RBC DHA and also more mature VEP acuity than did infants weaned to a control formula not containing DHA.79
N-3 LCPUFAs and Age-Related Macular Degeneration
DHA intake requirements related to vision are not well established after the first few years of life through middle age. During the aging process, age-related macular degeneration (AMD) becomes a major health issue and data suggest that DHA may be one of the dietary components that limit the progression of this disease. AMD is the third most common cause of blindness on a global basis80 and, in developed countries, is the leading cause of adult blindness.81 In the United States, the AMD prevalence of any severity is about 6.5% of the population above the age of 40 years and the prevalence of advanced AMD is 0.8%.82 AMD may present in one of two forms: central geographic atrophy (CGA, atrophy of the retinal pigment epithelium) or neovascular (NV, choroidal neovascularization) AMD. Advanced AMD is difficult to treat, and, therefore, prevention or treatment in its early stages would minimize the impact of the disease. Modification of life style, either through dietary changes or other means (e.g., smoking cessation) has the potential to reduce the substantial morbidity associated with AMD.
DHA is the predominant fatty acid of rod and cone outer segments and exceptionally prone to oxidation, a hallmark of AMD, due to its highly unsaturated character.83 It may at first appear counterintuitive that dietary n-3 LCPUFAs may have a beneficial role in preventing the initiation or progression of AMD. However, n-3 LCPUFAs may act via a number of mechanisms to reduce the potential for AMD.54 DHA and EPA are precursors of resolvins and neuroprotectins, agents that reduce the severity of inflammation and resolve the inflammatory process.84 These fatty acids modulate the expression of signal transduction genes and genes that code for proinflammatory cytokines.85,86 N-3 LCPUFAs and their metabolites exert cytoprotective effects in primary cell cultures of rat photoreceptors under conditions of oxidative stress87 and also have antiangiogenic and antivasoproliferative properties.54 Therefore, these fatty acids may alter the initiation or progression of AMD via numerous mechanisms.
Dietary Effects of N-3 LCPUFAs
Studies that target prevention of primary disease must be large in order to provide sufficient statistical power to evaluate contributing factors and must be of sufficient length to permit the disease to become apparent in a general population. In contrast, studies involving at-risk populations can involve fewer subjects, and studies in a population with active disease may be conducted for a shorter time. However, if the disease process is already underway, its progress may be difficult to stop or reverse by nutritional interventions. The effects of fish intake as a source of n-3 LCPUFA, as well as the consumption of EPA and DHA specifically, have been reported in both types of populations.
Epidemiologic studies which associates DHA and EPA consumption (or fish consumption) with AMD provide evidence for a protective role of n-3 LCPUFAs. A number of prospective studies have been reported.88,89,90,91,92,93,94,95 For example, AREDS3,88,89 enrolled individuals at moderate to high risk of developing AMD and followed them for 12 years. The size of this study (N = 1,837) and the elevated risk of developing AMD permitted assessment of both CGA and NV AMD development. Study participants in the highest quintiles for DHA, EPA, and DHA + EPA consumption were approximately 30% less likely to develop either CGA or NV AMD than were individuals in the lowest quintile. Results from Seddon et al. demonstrated that intake of fish,90 and specifically of DHA,91 was related to a reduced progression to advanced AMD. Studies involving individuals without risk factors for developing AMD have also demonstrated beneficial effects of n-3 LCPUFA consumption. Results from the Nurses’ Health Study and the Health Professionals Follow-up Study, the first published prospective study,92 evaluated a large population (42,743 women; 29,746 men) with no history of AMD for 12 years. DHA intake was related to a modest decrease in AMD, whereas linoleic acid intake was positively correlated with disease. The Blue Mountains Eye Study found that healthy individuals in the highest quintile of DHA and EPA consumption had a lower incidence of AMD than did individuals in the lowest quintile at 5 years.93 This study also found that fish (and n-3 LCPUFA) consumption was correlated with reduced incidence of AMD at 10 years.94 In a large study of women (N = 39,876) with no risk for development of AMD followed for 10 years, women in the top tertile for DHA or EPA consumption had a 35% to 40% reduction in the risk for development of AMD than did women in the lowest tertile.95 Women who consumed one or more fish meals per week were 42% less likely to develop AMD than women who consumed less than one fish meal per month, consistent with the results for DHA and EPA consumption.
The above data are suggestive of a beneficial relationship between dietary n-3 LCPUFAs and prevention of AMD; however, not all studies have been positive. In a cross-sectional study, Smith et al. reported that n-3 LCPUFA consumption may have a beneficial effect in advanced, but not in early ADM,96 whereas Chong et al. found a beneficial effect of consumption of n-3 fatty acids (the sum of ALA, EPA, and DHA), but not specifically EPA or DHA, in a prospective study.97 Although suggestive data are available concerning the beneficial effects of diets high in n-3 LCPUFAs, no studies reported to date have been randomized double-blind studies. The Age-Related Eye Disease Study II (AREDS II) is currently under way, comparing a treatment group (n-3 LCPUFAs, lutein, and zeaxanthin) to a placebo control group.4,98 Results of this double-blind, placebo-controlled trial will provide more definitive information concerning the role of n-3 LCPUFAs in AMD.
Only gold members can continue reading. Log In or Register to continue