Fig. 3.1
In our model for the protection of the DNA of CE cells from UV-induced damage the ROS, here H2O2, is produced by UV light. If free iron is present, this catalyzes the formation of hydroxyl radicals (OH·) by the Fenton reaction. And these hydroxyl radicals, being the most energetic ROS can damage DNA. If, however, nuclear ferritin is present to block the Fenton reaction, such damage will be prevented
Numerous studies have shown that DNA is a major target of UV-induced cellular damage. Damage to DNA can be direct, involving for example, DNA strand breakage and DNA–protein cross-linking. Or, it can be indirect, resulting from UV-induced formation of ROS, which, in turn, cause damage to DNA through alterations in the bases and deoxyribose moieties, strand breaks, and DNA cross-linking [21]. These changes can be mutagenic and carcinogenic [13]. ROS can also damage lipids and proteins. Thus, cells have evolved protective mechanisms against such oxidative damage [2] A growing body of evidence suggests that the ferritin-mediated sequestration of iron is one of these [3]—and that other iron-binding proteins, such as lactoferrin, can function similarly [42].
3.2 Free Iron and UV-Induced Damage to DNA
The deleterious effects of UV-induced ROS are greatly exacerbated by free iron, which catalyzes the Fenton reaction [46]. In the Fenton reaction (Fig. 3.1) Fe2+ catalyzes the conversion of hydrogen peroxide (H2O2) and superoxide (O2−) to the hydroxyl radical (OH·)—which, although acting over a short distance, is the most active ROS [28]. The Fenton reaction has been shown to increase H2O2-produced DNA breaks in preparations of nuclei [35,36] and rat hepatocytes [43]. At least part of the Fenton-reaction-mediated damage to nuclear DNA results from the presence of free iron in the nucleus [37]. The function of iron in the nucleus is unknown; however, its deleterious effects on DNA have been demonstrated [37]. It has also been determined that free iron can bind to specific sites on DNA and that this can generate Fenton-reaction-derived OH-radicals which specifically cleave the DNA at these sites [25,27,32].
Thus, we hypothesize that it is important to keep free iron in the nucleus at a low level—especially true for CE cells which are constantly exposed to environmental ROS-generating UV light and O2. Our studies suggest that one mechanism through which this is achieved is by having ferritin in the nucleus where it would be most effective in sequestering free iron.
3.3 Cytoplasmic Ferritin
Cytoplasmic ferritin has been extensively characterized. It is a highly conserved protein composed of 24 subunits (Fig. 3.2) with an aggregate molecular weight of >450 kD. It has a central core that can sequester as many as 4,500 molecules of iron in the ferric state. Each ferritin subunit (Fig. 3.2) has a molecular weight of ~19 kD and is comprised of five α-helices connected by loops of varying lengths [22]. In most mammalian species ferritin has heavy (H) and light (L) chain subunits, however, in the chicken, the species we have utilized for our work, only the H-chain has been detected—suggesting that the molecule is homopolymeric.
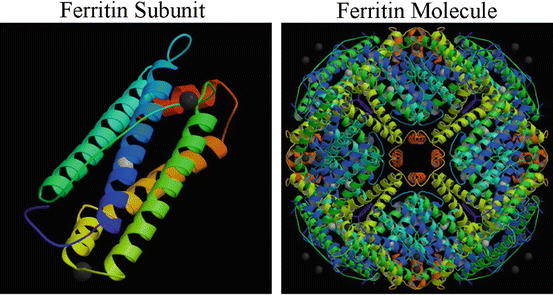
Fig. 3.2
Ferritin is a ubiquitous iron-storage molecule that is capable of sequestering up to 4,500 atoms of iron in its core. Ferritin subunits are comprised of five helical domains, and the “typical” ferritin molecule is a supramolecular complex comprised of 24 of these subunits
A number of the characteristics of the cytoplasmic ferritin molecule have been elucidated. This includes its supramolecular assembly, which involves interactions of the ferritin subunits, through specific molecular regions and sites in their primary sequence. To generate this complex, individual ferritin subunits associate with one another through interactions of their helical domains—which is a concept that is part of our proposed mechanism for the nuclear transport of ferritin (described later).
Assembly starts with the formation of dimers and ends as the 24-mer complex [29]. Functionally, molecular assembly is required for the importation of free iron and its sequestration within the core of the molecule—which removes free iron from participation in the generation of Fenton reaction-derived OH. radicals (Fig. 3.1) [17].
3.4 The Nuclear Ferritin of Corneal Epithelial Cells
In most cells ferritin is cytoplasmic—presumably due to the large size of the assembled molecule preventing its passive diffusion into the nucleus. However, our studies show that in avian CE cells, the ferritin is largely nuclear.1
The initial observation suggesting the possibility of ferritin being a nuclear component of CE cells arose serendipitously while screening—by IHC—a “whole-corneal” monoclonal antibody library for antibodies of potential interest for studying corneal development [49]. One of these antibodies (6D11), when tested on sections of anterior eyes, showed staining only of the CE. And subsequent, more extensive and detailed analyses using this antibody [10] showed its IHC staining to be localized exclusively to the nuclei of the CE cells (Figs. 3.3 and 3.12). Also, when this antibody was used to immunoscreen a cDNA expression library—constructed with cDNAs from chicken CE tissue—all of the positive clones had inserts whose sequence was “typical” cytoplasmic ferritin.
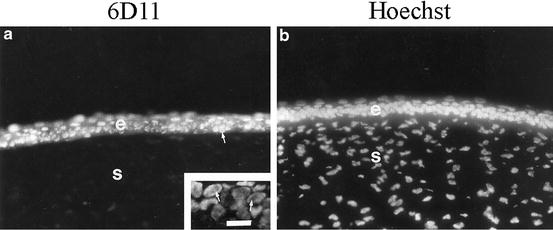
Fig. 3.3
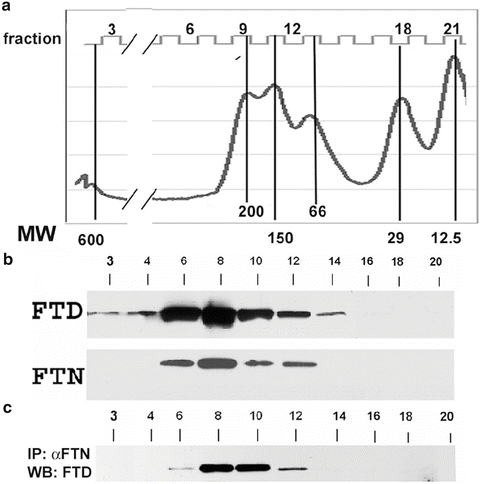
(a) Fluorescence IHC for ferritin on a section of mature, embryonic cornea. The CE is labeled “e”, the stroma is labeled “s”. The inset shows a higher magnification of the CE. The only detectible antibody staining is within the CE cell nuclei. There it is distributed throughout—except for the nucleoli (see inset). (b) Hoechst dye that identifies nuclei. This research was originally published in the Journal of Biological Chemistry; Cai CX, Birk DE, Linsenmayer TF. Ferritin Is a Developmentally Regulated Nuclear Protein of Avian Corneal Epithelial Cells. 1997;272(19):12831–12839, © The American Society for Biochemistry and Molecular Biology
Thus, avian CE cells have nuclear ferritin, and this is a “typical ferritin”, so it is not a different isoform that is modified to undergo nuclear transport. Instead, there is considerable evidence that CE cells have a tissue-specific nuclear transporter that associates with ferritin and carriers it into the nucleus. This transporter has been termed ferritoid—for its similarities to a ferritin subunit [38] and described later. In addition, evidence will be presented that following their nuclear transport, ferritin and ferritoid remain associated with one another as a unique, heteropolymeric complex(es) that has characteristics that differentiate it from all other known vertebrate ferritins [39].
3.5 The Role of Nuclear Ferritin in Protecting CE Cell DNA from Damage by UV Radiation
Based on the model presented in Fig. 3.1, studies were initiated to determine whether nuclear ferritin might be involved in preventing the DNA of CE cells from UV damage.
One approach was to UV-irradiate primary cultures of CE cells, and then evaluating the extent of strand breakage of their nuclear DNA—using fluorescence 3′-in situ-end labeling (ISEL) (as can be seen in Fig. 3.4).2
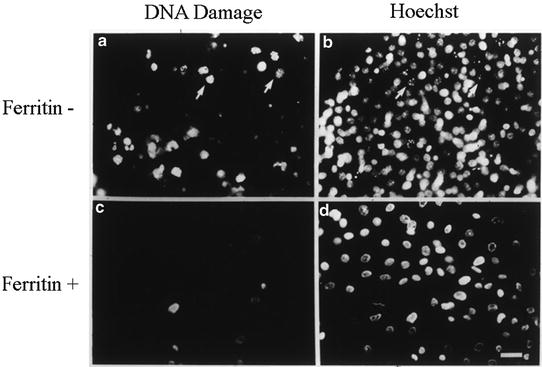
Fig. 3.4
All of our results are consistent with the nuclear ferritin of CE cells being protective against damage from UV irradiation and H2O2. In the experiment shown here, when CE cells are cultured in the presence of the iron chelator DFX (a, b), which prevents their synthesis of ferritin (ferritin−), and then are treated with UV light, extensive damage occurs (as determined by ISEL) when compared to cells with normal ferritin synthesis (c, d; ferritin+). Modified from [11]
This assay was used in a number of studies (e.g., [11], and reviewed in [30]). One study compared the relative effects of UV-induced damage to CE cells (with their nuclear ferritin)—versus cell types that do not have nuclear ferritin (e.g., corneal fibroblasts, and epidermal and dermal cells). Of these cell types, CE cells were by far the most resistant to DNA damage. Then, it was determined whether prevention of iron-mediated damage was involved—implicating the Fenton reaction. If so, the UV-induced damage should be exacerbated by elevated iron in the cell types without nuclear ferritin, but CE, with their normal elevated iron CE cells should be refractory to elevated iron. To test this, the effect of elevated iron on UV-induced damage was examined for CE cells versus corneal fibroblasts. For this, both types of cells were cultured in medium with the normal concentration of iron (normal-iron medium) versus medium in which the concentration of iron was elevated (high-iron medium). Then, the cells were irradiated and the damage was accessed by ISEL. In high-iron medium, the irradiated fibroblasts showed a large increase in damage to their DNA. However, for the CE cells increasing the iron produced little increase, if any, over their normal low level of damage.
These results are consistent with the CE cells having a mechanism(s) that prevents Fenton-reaction-mediated damage. They are also consistent with this mechanism involving ferritin in a nuclear location, as the nuclear ferritin of the CE cells was able to prevent UV-induced DNA damage—even in elevated iron—but the cytoplasmic ferritin of the fibroblasts did not confer protection.
Then to test directly whether the nuclear ferritin is responsible for preventing the UV-induced damage, experiments were performed in which the nuclear ferritin itself was manipulated. These experiments were based on the observations that pre-ferritin stage embryonic CE cells, when placed in culture, will initiate (precociously) the production of nuclear ferritin. However, if the iron chelator deferoxamine (DFX) is included in the medium, the initiation of ferritin synthesis is blocked (shown in detail in Fig. 3.15). Consistent with this nuclear ferritin being responsible for conferring protection, when the cultures with nuclear ferritin were irradiated (Fig. 3.4, ferritin+) the only cells showing an ISEL signal were occasional small rounded cells that were most likely naturally occurring, apoptotic cells—as these were also seen in un-irradiated cultures. However, in the cultures in which the nuclear synthesis was blocked (Fig. 3.4, ferritin−) most cells showed strong ISEL signals.
A modification of this experimental approach is based on the observation that the DFX blockage of nuclear ferritin synthesis can be reversed by replacing the DFX-containing medium with iron-containing medium (without DFX) (shown in Fig. 3.16). In addition, during the recovery from DFX there is a “window in time” in which some cells have initiated the production of nuclear ferritin, whereas others have not. Thus, irradiating cultures during this period makes it possible, in the same culture, to compare UV-induced damage to cells that have nuclear ferritin with ones that do not (data not presented, but see [11] and the review by Linsenmayer et al. [30]. Again, the cells that had nuclear ferritin (identified by IHC) showed little damage (by ISEL), and those that lacked nuclear ferritin showed damage. Also in the small population of cells that exhibited signals for both nuclear ferritin and damage to DNA, the ones with the least ferritin showed the most damage to DNA, and vice versa.
3.5.1 Protection from Damage by Reactive Oxygen Species (ROS)
If the protection afforded by CE nuclear ferritin involves the prevention of Fenton reaction-derived OH. radicals, protection should also be conferred against damage by ROS (e.g., H2O2) (as diagrammed in Fig. 3.1). So this was examined [9].
Similar to the UV-induced damage to DNA, the damage induced by H2O2 (determined by ISEL) was concentration and time-dependent. Of the four cell types tested (CE cells, corneal stromal cells, epidermal cells, and dermal cells), at all concentrations of H2O2, CE cells by far had the least damage. Also, CE cells showed a delayed temporal response to the appearance of damage, as compared to the other cell types.
And again, a protective role for nuclear ferritin was observed when DFX was used to inhibit the appearance of nuclear ferritin—followed by exposure to H2O2. Also consistent with the protection being effected through iron sequestration, for CE cells elevating iron in the medium did not increase damage by H2O2, whereas for epidermal cells it did.
It is also known that CE cells have other mechanisms that potentially may afford protection from oxidative damage—such as enzymatic scavenge pathways that exist for degrading H2O2 (e.g., catalase and glutathione peroxidase). So, in these studies it was also determined whether the protection afforded by nuclear ferritin is independent of these scavenge pathways. For these studies, the uptake and clearance of H2O2 was determined in CE cells versus several other cell types. This involved Fluorescence Activated Cell Sorting of cells that had been pre-labeled with an H2O2-sensitive dye and then were exposed to H2O2. Of the cell types examined, CE cells not only showed the highest uptake of the H2O2, but also the most rapid clearance. This clearance was most likely due to catalase and/or glutathione peroxidase—which is consistent with CE cells having scavenger enzymes that may have a role in protection against oxidative damage [12,19,34].
However, when H2O2-mediated damage was evaluated in the presence and absence of inhibitors of catalase and glutathione peroxidase there was no detectible difference in the damage to DNA. Thus, the protection afforded by nuclear ferritin is independent of cytoplasmic detoxifying enzymes.
3.6 Ferritoid: The Nuclear Transporter for CE Cell Ferritin
3.6.1 Isolation of a cDNA for Ferritoid and its Tissue Specificity
Ferritoid was initially detected as a “ferritin-like” component in a cDNA library of restriction enzyme-fragments of “up-regulated” corneal genes—using the subtractive hybridization procedure of [48].
As it was known from studies by others [18,41] that ferritin subunits associate with one another during the self-assembly of a supramolecular ferritin complex, we pursued the possibility that this clone might encode part of a ferritin-binding domain of the hypothesized nuclear transporter. If so, the encoded molecule should be specific for the CE—which was the next property that was examined and confirmed—both by in situ hybridization (Fig. 3.5a, b) and by IHC (Fig. 3.5c) using a ferritoid antibody.
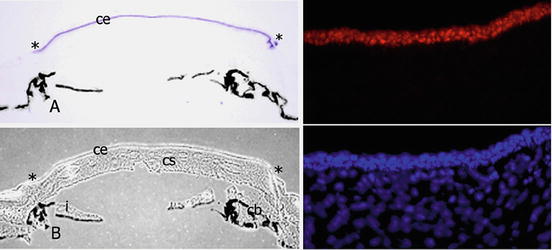
Fig. 3.5
If the tissue specificity of the nuclear ferritin transport is due to ferritoid, then ferritoid itself should be tissue-specific. As can be see in this figure, at both the mRNA and protein levels, ferritoid is specific for the CE. In (a) the in situ signal for ferritoid (purple) is found only in the CE—ending at the corneal-scleral junction (asterisks). b is a light micrograph of a and shows that the black regions in a and b are pigmented epithelia. The top right panel is IHC for ferritoid protein (red), and the bottom right panel is stained with Hoescht (blue). This research was originally published in the Journal of Biological Chemistry. John M. Millholland, John M. Fitch, Cindy X. Cai, Eileen P. Gibney, Kelly E. Beazley and Thomas F. Linsenmayer. Ferritoid, a Tissue-specific Nuclear Transport Protein for Ferritin in Corneal Epithelial Cells. 1997;278:23963–23970. © The American Society for Biochemistry and Molecular Biology
To obtain a full-length ferritoid cDNA, ferritin-like restriction-fragment clones from the subtracted library were used to screen another cDNA library, constructed using much longer cDNAs from chicken CE tissue. This screening identified other clones. The largest—diagrammed in Fig. 3.6—is likely to be full-length as it contains an uninterrupted reading frame (ORF) flanked by a 5′ consensus translation start site and a 3′ poly-A tail. Also, its conceptual translation product (shown in Fig. 3.6) contains two regions that fit with its proposed role as the nuclear transporter for ferritin.
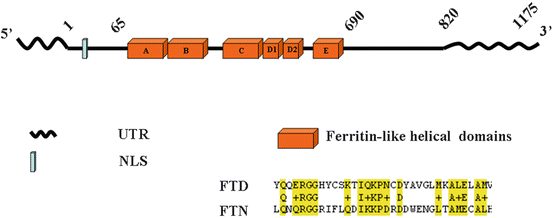
Fig. 3.6
The predicted structure of ferritoid includes: (1) a “ferritin-like” region that consists of all five helical domains of a ferritin, (2) a nuclear localization signal (NLS), and (3), 5′ and 3′ untranslated regions (UTR). Also shown in this figure is a conceptual amino acid sequence of a portion of one of the helical domains of ferritoid (FTD), and its comparison to the corresponding sequence in ferritin (FTN). The amino acids that are conserved between FTD and FTN are highlighted in yellow. From [38]
The first of these regions—which we propose is involved in the interaction/binding between ferritoid–ferritin—is “ferritin-like”. Consistent with this possibility, its conceptual amino acid sequence has 55 % similarity to ferritin, and it has most of the amino acids that studies by others [24] have shown in ferritin to be involved in the assembly of the supramolecular ferritin complex. Also, computer modeling [1,40] confirmed this region to have all five helical domains that are characteristic of a ferritin subunit (Fig. 3.6).
However, in ferritoid these helical domains are unique, as they are comprised of alternating short amino acid sequences that are highly conserved between ferritoid and ferritin (in Fig. 3.6 highlighted in yellow), and intervening among these sequences are equally short ones that have no similarity. The unique nature of this arrangement was extended by 3-D structural modeling (Fig. 3.7), in which the sequence of the ORF of ferritoid was inserted into the sequence of the known crystal structure of the H-chain subunit of human ferritin. This showed in ferritoid that the short “ferritin-like” amino acid sequences (shown in red in Fig. 3.7) are not distributed randomly along the lengths of the helices. Instead they are located preferentially along the outer surface of the helical domains—where we propose they interact with/bind to a ferritin subunit.

Fig. 3.7
In this figure, the crystal structure is of human ferritin-H is shown in four different planes. Into this structure, the deduced amino acid sequence of ferritoid has been superimposed. Also the amino acids of ferritoid that are conserved with those of ferritin are colored red, and the dissimilar amino acids are colored blue. As can be seen, in ferritoid, the conserved amino acids are preferentially located along the outer faces of the helices—where they would be available for interactions with ferritin. This research was originally published in the Journal of Biological Chemistry. John M. Millholland, John M. Fitch, Cindy X. Cai, Eileen P. Gibney, Kelly E. Beazley and Thomas F. Linsenmayer. Ferritoid, a Tissue-specific Nuclear Transport Protein for Ferritin in Corneal Epithelial Cells. 1997;278:23963–23970. © The American Society for Biochemistry and Molecular Biology
The second functional region—which is located just upstream of the ferritin-like region—contains a consensus SV40-like nuclear localization signal (Fig. 3.6, NLS) that experimental evidence shows is necessary for the nuclear transport of both ferritoid and ferritin (described next).
3.6.2 Functional Analysis of the Nuclear Transport of Ferritin by Ferritoid
To examine functionally whether ferritoid can transport ferritin into the nucleus, co-transfections were performed with epitope-tagged constructs for ferritoid (V5-tagged) and ferritin (myc-tagged) and the results were analyzed by double-labeled fluorescence IHC for the epitope tags.3 For these experiments, Cos-1 cells were used, as this is a cell type that had been used previously for transfection studies of ferritin [15]. In co-cultures transfected with full-length constructs for ferritin and ferritoid, in the cells that have both molecules, they are both exclusively nuclear [Fig. 3.8, NLS(+)]. Consistent with the NLS of ferritoid being functional and required, when the co-transfections were performed with the ferritoid deletion construct from which the NLS had been removed [Fig. 3.8, NLS(−)], the ferritin and ferritoid are distributed uniformly throughout the cell.4
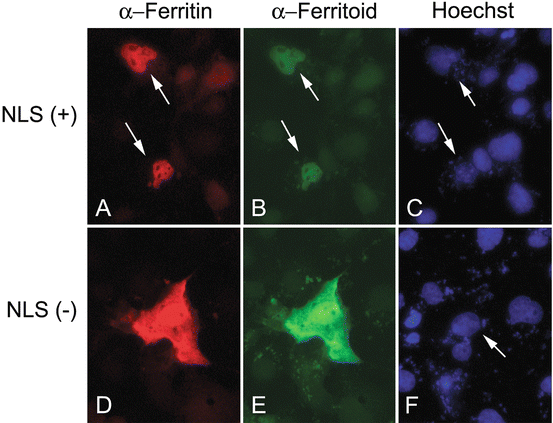
Fig. 3.8
COS-1 cells were co-transfected with a construct for ferritin, and constructs for ferritoid that are either full-length [with an intact NLS(+)] or ones from which the NLS has been deleted [NLS(−)]. The cells were analyzed by double-label IHC for ferritin (red) and ferritoid (green). Hoechst dye is also shown (blue). When the ferritin is co-transfected with the full-length ferritoid construct (NLS+), both ferritin and ferritoid are co-localized within the nucleus (arrows). When the co-transfections are done with the ferritoid construct that lacks the NLS(−) the ferritoid and ferritin are distributed uniformly throughout the cell. Modified from research originally published in the Journal of Biological Chemistry. John M. Millholland, John M. Fitch, Cindy X. Cai, Eileen P. Gibney, Kelly E. Beazley and Thomas F. Linsenmayer. Ferritoid, a Tissue-specific Nuclear Transport Protein for Ferritin in Corneal Epithelial Cells. 1997;278:23963–23970. © The American Society for Biochemistry and Molecular Biology
Then, to examine directly the association between ferritoid and ferritin, two approaches were employed. One was co-immunoprecipitation, and the other was a yeast two-hybrid binding assay.
The co-immunoprecipitations were performed on lysates of CE cells that had been co-transfected with epitope-tagged, full-length constructs for ferritoid (V5) and ferritin (myc). Then, the immunoprecipitates were analyzed by immunoprecipitation with antibodies for the epitope tags. The results showed that immunoprecipitates with either antibody contained both ferritoid and ferritin. Thus, ferritin and ferritoid associate with one another with an affinity strong enough to undergo co-immunoprecipitation.
For the yeast two-hybrid assays, the system employed not only shows qualitatively whether components interact with one another—as determined by the activation of a gene that is required for growth of a reporter yeast strain—it also provides quantitative information on relative strengths of these interactions. This latter parameter is based on the studies of [7] which showed that the stronger the interaction between two components, the greater the “relative plating efficiency” (RPE)—with RPE being defined as the number of colonies formed on medium that requires the interaction for growth, versus the number of colonies formed by a strain that grows constitutively.
For the ferritoid–ferritin analyses, the RPE used for normalization was the homopolymeric ferritin–ferritin interaction that occurs during assembly of a ferritin supramolecular complex. This was assigned a value of 100 % (Fig. 3.9). Then, the heteropolymeric ferritoid–ferritin interactions were evaluated, which gave an RPE of ~70 %. This result confirmed that ferritoid and ferritin interact with one another, and it also suggested that this heteropolymeric interaction is somewhat weaker than that which occurs between ferritin monomers. However, this heteropolymeric interaction is strong enough to produce complexes that within the nuclei of CE cells in situ are stable (as described next).
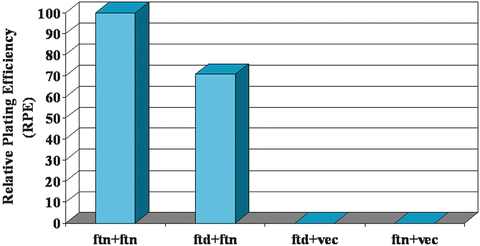
Fig. 3.9
The relative plating efficiencies (RPEs) of a yeast two-hybrid assay in which of ferritoid–ferritin interactions (ftd–ftn) are compared to ferritin–ferritin interactions (ftn–ftn; which are assigned a value of 100 %) shows that the ferritoid–ferritin interaction is slightly weaker—giving an RPE of ~70 %. In this assay, neither the ferritoid nor the ferritin, when inserted singly into the activation domain-containing vector, showed detectible growth
3.6.3 CE Nuclear Ferritin: A Stable Heteropolymeric Ferritin–Ferritoid Complex with Unique Properties
The studies already presented show that (1) ferritin and ferritoid interact with one another, and (2) this interaction is required for the nuclear transport of ferritin. However, these observations raised a number of additional questions concerning the ferritin and ferritoid that are present within CE cells in situ: including (1) the type(s) of complexes that are formed between these two components, (2) the subcellular localization(s) of these complexes, and (3) whether the complexes are transient—being present only during the transport process—or whether, once formed they remain as stable complexes. Also, if the complex(es) are stable, do they have unique characteristics/properties that distinguish them from other ferritin complexes? Certain of these questions have now been addressed.
3.6.4 Size(s) of the Ferritin–Ferritoid Complex(es)
For an initial characterization of the endogenous ferritin–ferritoid complexes of CE cells, extracts of CE tissue (from mature embryos) were fractionated by FPLC size-exclusion chromatography, followed by identification of the ferritoid and ferritin by Western blot.5 Using this approach, the fractions with ferritoid (FTD) eluted as a single peak (Fig. 3.10b), which by comparison with molecular weight markers (Fig. 3.10a), had a median molecular weight ~260 kDa (fraction #8). This is approximately half the size of a “typical” ferritin which has a molecular weight of ~470 kDa and is a 24-mer. Considerably lesser amounts of ferritoid were present in the highest molecular weight fractions (indicative of a “typical” 24-mer ferritin) and in lower molecular weight fractions that most likely are precursors of the higher molecular weight complexes (~50–100 kDa).