Cornea and Sclera
Daniel G. Dawson
Mitchell A. Watsky
Dayle H. Geroski
Henry F. Edelhauser
CORNEA
The cornea covers the anterior one-sixth of the total circumference of the globe (Fig. 1), whereas the sclera covers the remaining five-sixths. The cornea is a clear, transparent, colorless avascular structure richly supplied with sensory nerve endings that generally subserve touch and pain. There are no lymph vessels or other channels for bulk fluid flow. The interface between the corneal tear film and the ambient atmosphere provides roughly two-thirds of the refractive power of the human eye. The cornea itself is resilient, yet is described as viscoelastic in its response to stretching forces.
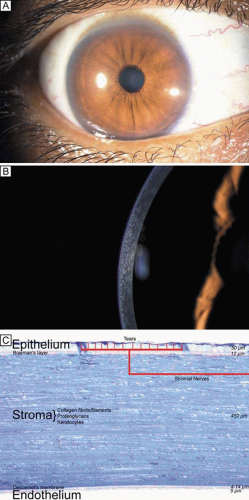 Figure 1. (A) Diffuse illumination slit-lamp view of the human cornea. (B) Slit-beam illumination slit-lamp view of the human cornea shows an optical section of the tissue. Notice the slight light scattering that occurs in the tissue, mainly from cellular components in cornea. (C) Histologic diagram of the major cellular and extracellular matrix components of the human cornea (toluidine blue 25×). |
Due to the avascular nature of the cornea, much of its oxygen requirement for metabolic activities comes from atmospheric oxygen dissolved in the tear film. When the eyelids are closed, oxygen can also predominantly enter the tear film from the superficial conjunctival capillaries. Most nutrients such as carbohydrates, vitamins, amino acids, and other substrates are primarily delivered to the cornea through the leaky corneal endothelial barrier with a minor contribution from the vascular arcades of the limbus. Certain growth factors, immune constituents, and other substrates like retinol are also secreted by the lacrimal gland and delivered to the cornea by the tears. Carbon dioxide and other metabolic end products are removed across the corneal endothelium to the aqueous humor, by the tear film, or through the limbal capillaries.
EMBRYOLOGY AND DEVELOPMENT
Following lens vesicle formation, which occurs between 4 to 5 weeks of gestation (27 to 36 days), surface ectodermal cells cover the defect left by lens vesicle invagination and become the primitive corneal epithelium composed initially of two cell layers. Similar to the avian cornea, the primitive epithelium of the primate cornea appears to be responsible for forming a prominent primary acellular corneal stroma, also known as the Bowman’s layer, which becomes detectable around 20 weeks of gestation. It should be noted, however, that the primate cornea’s ectodermal phase (primary acellular stroma) is de-emphasized compared to the mesodermal phase (secondary cellular stroma) and is less well organized in ultrastructure compared to that of the avian cornea.
Sometime between when the eyelids fuse together (8 weeks gestation) and by the time the eyelids open (26 weeks gestation), the epithelium stratifies to four to five cell layers thick. The basal epithelial cells, which secrete an underlying basement membrane called the basal lamina, are cuboidal to columnar. Above this layer are interdigitating wing cells and, most superficially, a layer of flattened squamous epithelial cells. Early in gestation, adhesion complexes (hemidesmosomes, anchoring fibrils, and anchoring plaques) on the basal surface of the epithelium are absent. Rudimentary adhesion complexes only become detectable by 19 weeks of gestation. With further development in utero, the number of hemidesmosomes increases, anchoring fibril penetration into the Bowman’s layer increases, and Bowman’s layer thickens.
A first wave of neural crest-derived mesodermal cells begins to extend beneath the corneal epithelial cells from the limbus around 5 weeks of gestation (33 days); these cells form the primitive endothelium. The primitive endothelium is initially composed of two cell layers, which by 8 weeks of gestation become a monolayer that starts to produce Descemet’s membrane. The epithelium and endothelium remain closely opposed until 7 weeks of gestation (49 days) when a second wave of mesoderm begins to grow centrally from the limbus between the epithelium and endothelium. This tissue forms the secondary cellular corneal stroma, and extracellular collagen is produced within a few days. During the third month of gestation, Descemet’s membrane is secreted by the endothelial cells and can be recognized in histologic sections. By the seventh month of gestation, the cornea resembles that of the adult in most structural characteristics other than size.
At birth in the full-term infant, the horizontal diameter of the cornea is only around 9.8 mm (surface area 102 mm2), or approximately 75% to 80% of the size of the adult human cornea. (Note that the posterior segment is less than 50% of adult size at birth.) Additionally, at birth, the cross-sectional thickness of the epithelium averages 50 μm, the Bowman’s layer averages 12 μm, the central cellular corneal stroma averages 450 μm, Descemet’s membrane averages 4 μm, and the endothelium averages 5-μm thick.
During infancy, the cornea continues to grow over the first years of life, reaching adult size at 2 years old with a horizontal diameter of 11.7 mm (surface area 138 mm2) and changes very little in size, shape, and optical properties thereafter. The only significant structure in the cornea that thickens with age is the Descemet’s membrane, which gradually increases an additional 6 to 11 μm from birth to senescence.
OPTICAL PROPERTIES
The average index of refraction of the cornea and tear film taken as a whole is about 1.376. The refractive power for the anterior surface of the cornea may be computed by the following formula:
D = n′-n/r = D = 1.376-1.000/0.0078 =48.2
where D equals diopters of optical power, n is the index of refraction of air (1.000), n′ is the index of refraction of the whole cornea, and r is the radius of curvature of the anterior corneal surface in meters.
The posterior surface of the cornea is bathed with aqueous humor. By applying the same formula, we arrive at the following value for the optical power of the posterior corneal surface, where n′ is the refractive index of aqueous humor (1.336):
D = n′ -n/r = 1.336-1.376/ 0.0065=-6.2
Therefore, the total optical power of the cornea is 48.2 – 6.2, or approximately 42.0 D, which is about two-thirds of the 60.0 D total optical power of the human eye. Because the cornea is thinner in the center than in the periphery, it should act as a minus lens but functions as a plus lens because the aqueous humor neutralizes most of the minus optical power on the posterior corneal surface. If we compute the power of the posterior corneal surface in air, we would find the following:
D=1.000-1.375/0.0065 =-96.0
By subtracting the plus power of the anterior surface, the resulting net power for the cornea in air becomes -96.0 D + 42.0 D = -54.0 D, a powerful minus lens.
From the previous calculations, it is obvious that the most important refracting surface is that of the anterior cornea. However, if a large air bubble is placed in the anterior chamber so that it contacts the corneal endothelium or if the anterior surface of the cornea is submerged in water—thereby destroying the corneal-air interface—tremendous changes in the refractive power of the eye occur. When the eye is open underwater, the optical imagery is extremely blurred; the index of refraction of water is quite similar to that of the cornea and most of the optical power of the anterior corneal surface is lost. If the tear film–air interface is maintained by the use of a mask or goggles, then underwater mask vision is as sharp and clear as normal terrestrial vision.
A contact lens placed on the corneal surface is an efficient way of correcting optical errors in the dioptic system of the eye. This is particularly true for errors that are caused by abnormalities of corneal curvature. The contact lens placed upon the corneal surface has the same index of refraction as the cornea and becomes covered immediately with the normal corneal tear film. Therefore, the contact lens becomes, in effect, a new part of the cornea.
Several surgical procedures have also been developed to permanently alter the curvature of the cornea, thereby reducing nonpathologic refractive errors. The procedures most commonly performed today include laser-assisted in situ keratomileusis (LASIK), photorefractive keratectomy (PRK), and astigmatic keratotomy (AK). LASIK and PRK are surgical techniques that use an excimer laser (193 nm) to ablate the anterior corneal stroma with submicron accuracy and create a new refractive surface. LASIK is different from PRK in that it involves a mechanical or laser created lamellar keratectomy step before the excimer ablation step, which is applied to the underlying residual mid-stromal bed. PRK ablation directly occurs on the surface of the cornea (Bowman’s layer and anterior stroma). Both procedures have been used on myopic and hyperopic eyes with up to a mild degree of astigmatism and produce similar visual outcomes. AK is an incisional keratotomy technique that cuts up to 90% depth arcs in the peripheral cornea using a scalpel to reduce astigmatism by flattening the cornea in the visual axis over the area of the cut. AK is usually performed in association with cataract surgery.
GROSS ANATOMY
When viewed anteriorly in the living eye, the adult human cornea is somewhat elliptical; the largest measurement is typically in the horizontal meridian (mean 11.7 mm) and the smallest is in the vertical meridian (mean 10.6 mm). The elliptical configuration is brought about by an anterior extension of the opaque scleral structure superiorly and inferiorly. When viewed from the posterior surface of the dissected eye, the cornea is circular, with an average horizontal and vertical measurement of 11.7 mm. The average radius of curvature of the anterior corneal surface is 7.8 mm, which is significantly less than the 11.5 mm average radius of curvature of the sclera. This results in a small, 1.5- to 2-mm wide transition zone that forms an external and internal surface groove, or scleral sulci, where the steeper cornea meets that of the flatter sclera. These sulcus typically are not as obvious to see clinically because they are filled in by overlying episclera and conjuctiva or trabecular meshwork.
The tissue in this transition zone is known as the limbus and is important because it is used as surgical landmark for various anterior segment surgeries, contains corneal epithelial stem cells, contains the conventional outflow pathway for the aqueous humor, is the inciting site of pathology in a few immunologic or cancerous diseases, and possibly contains stem-like corneal endothelial cells (Fig. 2). The limbus is a definite anatomic landmark, or geographical reference, used for planning surgical entry into the anterior segment because it appears clinically as a blue transition zone. Therefore, an incision placed anterior to blue zone is anatomically in the peripheral cornea, safely inside the trabecular meshwork.
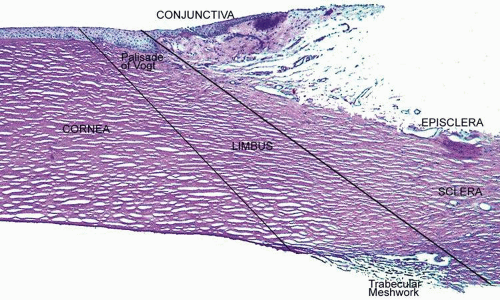 Figure 2. Photomicrograph demonstrating the relationship of the peripheral cornea, limbus, sclera, episclera, and conjunctiva (PAS, 20×). |
The cornea is thinner in the center, measuring on average 520 μm and increases in thickness to approximately 700 μm as it reaches the limbus.1,2,3,4 The central 4 mm of the cornea overlying the pupil contains the optical center of central vision. It typically has a regular surface and structure, and commonly is near exact in spherical configuration. The more peripheral portions of the cornea are often slightly irregular and torical in configuration rather than spherical, giving the whole cornea a hyperbolic shape. If the central cornea is not exactly spherical, the condition of astigmatism (stigma meaning a point) usually results. With astigmatism, an optical image without a single point focus forms within the eye resulting in two line foci.
The average radius of curvature for the posterior corneal surface is shorter than the anterior corneal surface at 6.5 mm. In adult humans, the conjunctival surface area has been measured at 17.65 cm2, and the corneal surface area measures 1.38 cm2, giving a c/c ratio of 12.8.5 Because topical lipophilic drug delivery to the anterior chamber occurs primarily through the cornea with the conjunctiva supplying a minor secondary delivery route, surface area ratios of the cornea and conjunctiva are important because a large conjunctiva-to-corneas (c/c) surface area ratio results in less drug delivery to the anterior chamber. Therefore, surface area ratios need to be considered when comparing drug delivery studies.
MICROSCOPIC ANATOMY, ULTRASTRUCTURE, AND PHYSIOLOGY OF THE EPITHELIUM
The anterior surface of the human cornea is covered by a transparent, nonkeratinized, stratified (five- to seven-cell layer) squamous epithelium uniformly around 50 μm in thickness that is continuous with the epithelium of the limbus and conjunctiva (Figs. 1, 2, and 3). The basal corneal epithelial cells actively secrete extracellular material (type IV collagen, laminin, heparin, and small amounts of fibronectin and fibrin) that forms an underlying 75-nm thick basement membrane called the basal lamina. On electron microscopy, the morphology of basal lamina appears to be composed of two distinct layers: a 25-nm thick lamina lucida and a 50 nm thick lamina densa (Fig. 3).
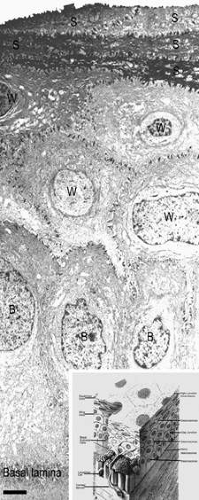 Figure 3. Transmission electron micrograph (3,500×) of the central corneal epithelium with a summary diagram (inset). Microvilli project from the anterior corneal surface into the tear film. S, squamous cells; W, wing cells; B, basal epithelial cells. Bar = 1 μm. (Inset modified from Hogan MJ, et al. Histology of the human eye. Philadelphia, WB Saunders, 1971.) |
The cytoplasm of a corneal epithelial cell primarily contains cytoskeletal intermediate filaments and has sparse cytoplasmic organelles (i.e., mitochondria, endoplasmic reticulum, and golgi apparatus). The predominant cytoplasmic filament is keratin, whereas actin and microtubules are two other major types found in corneal epithelial cells. The epithelial cells are held together to one another by numerous anchoring junctions called desmosomes, whereas the basal surface of the epithelium adheres to the basal lamina and underlying Bowman’s layer through aa adhesion complex composed of hemidesmosomes, anchoring fibrils (type VII collagen), and anchoring plaques (Fig. 4). The function of the corneal epithelium is twofold: (a) to form a barrier from the environment to the corneal stroma of the cornea, and (b) to form a smooth refractive surface on the cornea through interaction with the tear film.
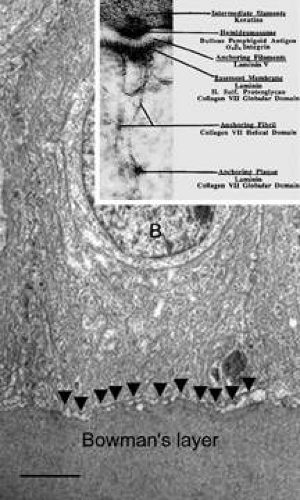 Figure 4. Transmission electron micrograph (10,000×) of a basal epithelial cell showing the adhesion complexes (arrowheads) that anchor it in place onto the Bowman’s layer and summary inset. B, basal epithelial cell. Bar = 1 μm. (Inset from Albert and Jakobiec: Principles and practice of ophthalmology. Philadelphia, WB Saunders, 2000.) |
The epithelial cells differentiate from the basal layer to form two to three cell layers of wing cells and finally to form two to three cell layers of squamous cells (Fig. 3). The squamous cells form a barrier junction because they are surrounded by a continuous encircling band of zonula occludens tight junctions, which serve as a semipermeable, high-resistance (12–16 kΩ cm2) membrane6,7 by closing off the intercellular space. This barrier prevents the movement of fluid from the tears into the stroma and also protects the cornea and intraocular structures from infectious pathogens. The apical surface of the corneal epithelium is specialized to maintain the tear film as microplicae and microvilli on the surface of the most superficial epithelial cells is covered with a glycocalyx and membrane-spanning mucins (MUC 1 and possibly MUC 4); altogether these structures and substances form the 1.0 μm thick mucinous layer of the tear film (Fig. 5).8,9,10
 Figure 5 Transmission electron micrograph (10,000×) of surface epithelium from a specimen specially-preserved (glutaraldehyde + cetylpyridium chloride) and stained (tannic acid) to show the mucinous layer of the tear film (glycocalyx + membrane-bound mucins). Inset is a summary diagram showing how the tear film layers interact with the microvillae of the surface squamous epithelial cells. S, squamous cells; W, wing cells. Arrowheads = zonula occludens tight junctions. Arrowheads = zonula adherens tight junctions. Bar = 1 μm. |
The tear film typically measures 7 to 10 μm in thickness and contains three layers (mucinous, aqueous, and lipid layers). Recently, the aqueous layer of the tear film has been found to contain gel-forming mucins (MUC 5AC and 2) along with lysosyme, immunoglobulin A, transferring, defensin, and trefoil factor (Fig. 5, inset).10 Overall, the tear film functions in maintaining a healthy ocular surface by preventing ocular infections and forming a smooth optical surface required for clear vision. Deficiencies in the components that compose any of these layers potentially can cause ocular surface disease.
The corneal epithelium is in a state of constant healing as squamous cell are continuously
shed into the tear film (Fig. 6). It is estimated that all the cell layers of the corneal epithelium completely turn over every 7 to 10 days. The epithelial surface is maintained by basal epithelial cells, which can undergo mitosis resulting in two daughter cells that are found anterior to the basal cell layer, thus forming two wing cells and eventually squamous cells.11,12,13 This delicate balance of shedding followed by proliferation is critical in maintaining a smooth and uniform epithelial surface around 50 μm in thickness. The shedding step is primarily induced by the friction that occurs from involuntary eyelid blinking, which happens on average every 7 seconds.
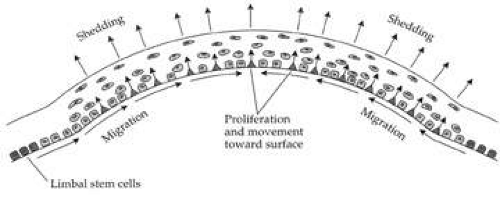 Figure 6. Diagram showing how the basal epithelial cells are continually replenished by a stem cell population that resides in the limbus. The basal epithelial cells migrate forward from the periphery to the center of the cornea. When they undergo mitosis, both daughter cells move into the anterior layers of the epithelium and continually move up in epithelium until eventually being shed. (From Thoft RA, et al. The X, Y, Z hypothesis of corneal epithelial maintenance. Invest Ophthalmol Vis Sci 24:1442, 1983.) |
The signal for basal epithelial cell proliferation probably comes via the gap junctions, especially in the more basal layers, with connexin 43 being one of the common gap junction proteins present.7,14 Interestingly, gap junction communication between corneal epithelial cells is promoted by low concentrations of bicarbonate, which is abundant in the aqueous humor and tear film.15 The barrier function of the epithelium itself is due to zonula occludens tight junctions between the most superficial epithelial junction, which are made up of the tight junction proteins ZO-1, occludin, and claudin-1 as well as other claudin sub-types.16 Under normal conditions, this barrier is extremely tight, preventing the movement of ions between the superficial squamous cells. In some instances (e.g., trauma, contact lens wear, infection), however, this barrier can be compromised. Activation of protein kinase C (PKC) by the mitogen-activated protein kinase (MAP kinase) signaling pathway, which in itself is activated by external stimuli such as growth factors and tyrosine kinases, appears to cause disruption of the epithelial barrier and perhaps is an initiating factor for basal cells to proliferate.17
In addition to basal epithelial cell mitosis, the corneal epithelium is maintained by migration of new basal epithelial cells into the cornea from the limbus. The cells migrate centripetally at about 120 μm/week and originate from a subpopulation of limbal epithelial cells.18,19,20 Therefore, it appears that the corneal epithelium is maintained by a balance among the processes of limbal cell proliferation, basal corneal epithelial cell migration and proliferation, and shedding of superficial squamous corneal epithelial cells.21,22 This concept has been coined the X, Y, Z hypothesis by Thoft [X (basal corneal epithelial cell proliferation) + Y (limbal cell proliferation and basal corneal epithelial cell centripetal migration) = Z (epithelial cell loss from the surface)].22
When this equilibrium is disturbed, corneal epithelial cell wound healing typically begins. Interestingly, after injury, these processes typically return back to equilibrium with a certain degree of adaptability. For instance, after epithelial and stromal injury, if a stromal deficit occurs (e.g., corneal ulcers) then the epithelium can still maintain a smooth anterior surface either by developing elongated hypertrophic basal epithelial cells and/or by developing epithelial hyperplasia (> six cell layers).23
As first described in the early 1960s by Maumenee, and expanded upon in the early 1970s by the work of Davanger and Evensen and in the early 1980s by Shapiro, Friend and Thoft, the limbal epithelium contains cells that have the ability to resurface the cornea following loss of the corneal epithelium.24,25,26 This concept was applied clinically by Thoft in 1977, who described the use of autologous limbal epithelial transplants to treat unilateral chemical injury.27 Subsequently, further study has determined that a portion of limbal basal epithelial cells are actually undifferentiated corneal epithelial cells, also known as corneal epithelial stem cells.28,29 These stem cells have the characteristics of being extremely long-lived, with very slow cell cycle times and a high proliferative potential. They anatomically are found within a well-defined, protective microenvironment called the palisades of Vogt (Fig. 7) and are controlled via delicate regulatory mechanisms.30,31,32 The progeny of stem cells are referred to as transient amplifying (TA) cells, which are cells that have more rapid cell cycles, yet have far less replicative potential than their parent stem cells. During terminal differentiation from the basalepithelial cell layer, the daughter cells migrate anteriorly in the epithelium becoming wing cells and, lastly, squamous cells.
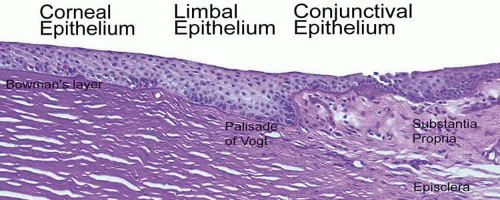 Figure 7. Photomicrograph of a palisade of Vogt, which is where the epithelial stem cells are thought to reside (PAS, 100×). |
When the mammalian cornea is mounted in an Ussing chamber, it generates a transepithelial potential of 25 to 40 mV. This relatively high voltage is consistent with the low ionic conductance of the apical epithelial cell membranes and the high resistance of the tight junctions of the paracellular pathway, which normally is in the range of 12 to 16 kΩ cm2.6 Almost 50% of the short-circuit current across the corneal epithelium is carried by chloride ions moving through apical membrane channels into the tears. This current is due to ionic gradients set up by epithelial transport of Na+ and Cl– ions. Na+ ions are pumped from the epithelial cells toward the stroma by the Na+/K+ adenosine triphosphatase (ATPase) in the lateral membranes of the cells, which, as in other cells, maintains an inward Na+ gradient. A Na+-chloride co-transporter, also located in the lateral membrane, facilitates the influx of Na+ down its electrochemical gradient, carrying along chloride ions. The chloride ions then diffuse through channels in the apical membranes. This chloride secretion is blocked when the Na+/K+ ATPase is inhibited by ouabain, demonstrating the coupling of chloride secretion to Na+ transport.33,34,35,36
In addition to the transport mechanisms described, the corneal epithelial cells also contain a Na+/H+ exchanger and a lactate-H+ co-transporter. These transport mechanisms serve to regulate intracellular pH by extrusion of lactate and H+ ions.37 The above discussion raises questions concerning the overall role of these transport mechanisms and second-messenger systems in corneal epithelial function. It has been demonstrated in vitro that ion transport can osmotically move water from the stroma to tears6; however, in vivo epithelial ion transport probably has a limited role, if any, in corneal deturgescence as compared to the endothelium.
Finally, it has been shown in both animal and human specimens that the corneal epithelium, although devoid of melanocytes, does contain immune cells. The basal epithelial cell layer of the peripheral cornea, along with the limbus and the conjunctiva, appears to have a subpopulation of cells intermixed that are bone-marrow–derived immune surveillance cells with high constitutive expression of major histocompatibility complex (MHC) type II antigen and co-stimulatory molecules.38 This type of immune cell has previously been termed a Langerhans cell,39 which functions as a “professional” antigen-presenting cell with an extraordinary capacity to initiate T-cell lymphocyte-dependent responses.40 The functional steps of this cell type include the uptake and processing of antigens, the migration out of the cornea to lymph nodes where they stimulate naïve T-lymphocyte immune responses by presenting antigens and overexpressing co-stimulatory molecules. Recently, the central corneal epithelium also was found to have a similar subpopulation of immune cells that are apparently “immature” Langerhans cell types because their constitutive expression of MHC type II antigen and costimulatory molecules is low.41 Interestingly, these “immature” Langerhans cells under certain circumstances (e.g., inflammation or trauma) may develop the requisite signals for T-cell priming.41
MICROSCOPIC ANATOMY, ULTRASTRUCTURE AND PHYSIOLOGY OF THE STROMA
The corneal stroma accounts for 90% of the corneal thickness. It is predominantly composed of water (78% hydrated or 3.5 g H2O/g dry weight), which is stabilized by an organized structural network of insoluble and soluble cellular and extracellular proteins (Fig. 1).42 The dry weight of the adult human corneal stroma is made up of collagen (68%), keratocyte constituents (10%), proteoglycans (9%), and salts, glycoproteins, or other substances.43
Collagen is a water-insoluble structural protein organized into an inextensible scaffold that forms the basic structural framework of connective tissues. In the human cornea, stromal collagens are poorly-ordered in the Bowman’s layer (i.e., the primary acellular stroma that is presumably ectodermally-derived), whereas they form a highly-ordered lattice-like structure in the secondary cellular stroma (neural crest-derived).44 Overall, the corneal collagens are functionally important in establishing tissue transparency and in resisting tensile forces because collagen fibrils and filaments hold the cornea together (i.e., cohesive strength), ultimately defining the size of the tissue.
Keratocyte components make up the second major component of the cornea’s dry weight. Interspersed between collagen lamellae throughout the secondary cellular stroma, keratocytes form a closed, highly-organized syncytium. Keratocytes are neural crest-derived cells that enter the cornea during the second mesenchymal wave of corneal embryologic development. They function as modified fibroblasts during neonatal life, forming the extracellular matrix of the secondary cellular stroma. Subsequently, they typically remain in the cellular stroma throughout life as modified fibrocytes where they maintain the extracellular matrix of the corneal stroma, usually in an inconspicuous, or relatively transparent, fashion.
Proteoglycans, which are water-soluble glycoproteins, are the third major component of the cornea’s dry weight. Each proteoglycan molecule consists of a protein core with a covalently attached anionic polysaccharide side chain called a glycosaminoglycan (GAG). The proteoglycans are three-dimensionally ordered in the corneal stroma because the protein core of the molecule is noncovalently attached to collagen fibrils evenly throughout the tissue, whereas the GAG sidechain extends into the interfibrillar space where it acts as a pressure-exerting polyelectrolyte gel.45,46,47 After Hedbys showed that the cornea collapses to around 20% of its volume if the proteoglycans in the tissue are precipitated with cetylpyridinium,48 it has become quite apparent that the primary function of proteoglycans is to provide tissue volume, maintain spatial order of collagen fibrils, and resist compressive forces. Water, collagens, proteoglycans, and keratocytes work together to maintain and establish a transparent cornea, while also creating a tough and resistant structure that keeps ocular integrity intact and maintains a stable shape.
Interestingly, although the cornea primarily absorbs light between 200 to 295 nm (ultraviolet)49 and transmits variable amounts of light from around 300 nm in wavelength (ultraviolet) to 2,500 nm in wavelength (infrared)50, peak transmission (95% to 99% of the incident light) occurs along wavelengths where light is visible to humans (visible spectrum of light = 400 to 700 nm). The remaining portion that is not directly transmitted (1% to 5%) is scattered in all directions by the cornea in a wavelength dependent fashion or absorbed by the tissue (Fig. 8).51 Clinical slit-lamp examination and in vivo confocal microscopy suggests that most of the scattered light (i.e., reflected and scattered light) comes from cellular components in the cornea (endothelial cells > epithelial cells > nerve cells > keratocytes) as opposed to collagen fibrils (Figs. 1B and 8, bottom).52,53
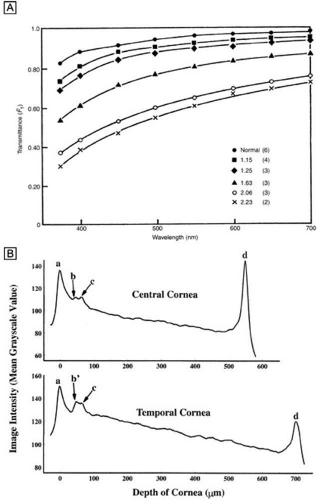 Figure 8. (A) Experimental values for the fraction of light transmitted through normal and edematous rabbit corneas as a function of wavelength. The ratio of the thickness of the edematous corneas to normal thickness values and the number of corneas used for each curve are given in the key. (From Farrell RA, et al. Wave-length dependencies of light scattering in normal and cold swollen rabbit corneas and their structural implications. J Physiol233:589, 1973.) (B) In vivo confocal microscopy back-scattered light intensity profiles from the central and temporal portions of a 25-year-old human cornea. Intensity peaks correspond to the (a) epithelium, (b) subbasal nerve plexus, (c) most anterior keratocytes layer, and (d) endothelium. (From Patel SV, et al. Normal human keratocyte density and corneal thickness measurement by using confocal microscopy in vivo. Invest Ophthalmol Vis Sci 42:333, 2001.) |
Collagen molecules measure 1.5 nm in width by 300 nm in length and are composed of a triple helix of three alpha chains (Fig. 9, top). There currently are 19 known collagen types as determined by the combination of alpha-chains that form the collagen molecule. The most common types usually aggregate into structural, banded (repeating pattern of polar amino acids seen on transmission electron microscopy [TEM]) fibrils by ordering themselves into a quarter-staggered parallel arrangement that is further stabilized in position by covalent intermolecular cross-links (Fig. 9, middle).
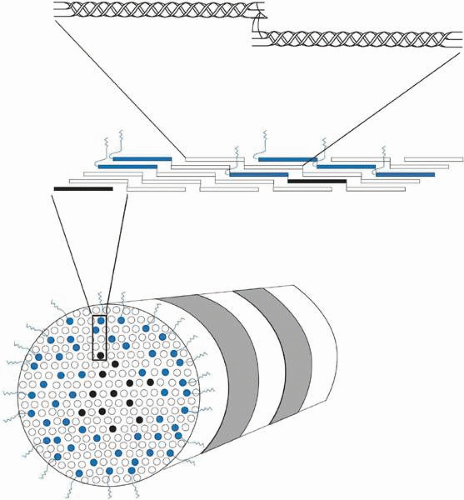 Figure 9. Cross-sectional oblique view of a 25-nm diameter, heterotypic, banded (periodicity = 68 nm) corneal stromal collagen fibril (bottom) composed of type I (white), III (black), and V (blue) collagen molecules. The amino-terminal domains on the type V collagen molecules appear to be important in regulating collagen fibril diameter as they project external to the fibril surface and presumably block further accretion of collagen molecules through steric and/or electrostatic hindrance effects. Notice that the collagen molecules on longitudinal view (middle) are aligned in a parallel, quarter-staggered (68 nm) arrangement with 40 nm gaps between molecules. Also, note that the longitudinal view (top) shows clearly that the ends of the alpha chains in each collagen molecules form intermolecular cross-links with adjacent collagen molecules. |
Collagen fibrils are generally heterotypic (composed of two or more types of collagen molecules) and reach certain diameters based on their composition of collagen types. The fibrils in the corneal stroma are a co-polymerization of collagen types I, III, and V molecules that form uniform 22-nm diameter fibrils in the acellular Bowman’s layer and uniform 25-nm diameter fibrils in the cellular corneal stroma with only slight variability (Figs. 9, bottom and 10).54,55 Although the refractive index of collagen fibrils (1.47) is different from that of the extrafibrillar matrix (1.35), the highly uniform size (25 ± 2 SD nm) and interfibrillar spaces (40 ± 5 SD nm) along with the predominantly parallel directionality of these fibrils results in a highly-ordered lattice-like array of fibrils (i.e., not a true crystalline lattice, but one with short-range order) that allows transparency of the cornea due to destructive interference effects (Fig. 10B, C).56
 Figure 10. (A) Low magnification (4,750×) TEM of predominantly orthogonally stacked lamellae in the middle third of cellular corneal stroma. (B) Higher magnification (72,500×) TEM of two lamellae in the middle third of cellular corneal stroma. One lamellae is in longitudinal view (top) and other is in cross-sectional view (bottom). Notice the uniformly 25-nm diameter collagen fibrils and the 40-nm diameter interfibrillar spaces that demonstrate only a short-range order (i.e., slight variability), but do not form a true crystalline lattice. (C) Cross-sectional diagram of collagen fibrils arranged in a true crystalline lattice arrangement. Size of a wavelength of light is shown above for comparison. Bars = 1 μm. (From Maurice DM. The structure and transparency of the cornea. J Physiol 136:263, 1957.) |
Collagen type VI is the third most common type of collagen in the corneal stroma, but is unique in that it is only able to aggregate into repeating tetramers of type VI molecules. Thus, it forms only 10 to 15 nm diameter, beaded (20 × 30 nm diameter ovals with a periodicity of 100 nm), nonbanded filaments (Fig. 11A). Functionally, it acts as a bridging filament that binds corneal lamellae together where they cross each other (Fig. 11B, C). Along with fibril-associated collagens with interrupted triple helices (FACIT collagens, type XII and XIV collagen molecules), it also bridges intralamellar fibrils together (Fig. 11D).57,58 Overall, this three-dimensional, supramolecular scaffold created by human corneal stromal collagens results in a one-dimensional ordered (∼22 nm diameter collagen fibrils; ∼ 40 nm interfibrillar spaces × random directionality) 12-μm thick acellular Bowman’s layer (Figs. 12A and 13A) and a three-dimensionally-ordered (∼25 nm diameter collagen fibrils; ∼ 40 interfibrillar spaces × parallel directionality) series of successive stacks of lamellae in the cellular stroma measuring in the center of cornea around 450 μm in thickness.59,60,61
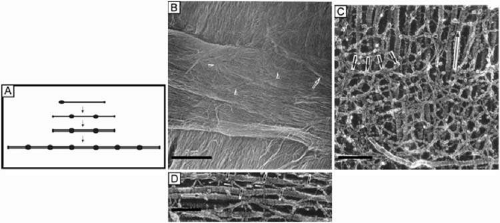 Figure 11. (A) Diagram of a type VI collagen molecule and how it assembles into filaments by aggregating into repeating tetramers of type VI molecules (periodicity of 100 nm). (B) Low magnification (6,200×) SEM showing bundles of collagen filaments extending between lamellae (arrow) and a loose meshwork of collagen filaments on the posterior surface of a corneal lamellae where an adjacent lamellae crosses it (arrowheads). Bar = 5 μm. (From Komai Y, et al. The three-dimensional organization of collagen fibrils in the human cornea and sclera. Invest Ophthalmol Vis Sci 32:2244, 1991.) (C) High magnification (115,000×) quick-freeze, deep-etched electron micrograph showing a loose meshwork of interlamellar beaded filaments with a periodicity of 100 nm (thick arrows) which appear to bind to collagen fibrils (long arrow) by their beads (arrowhead) and join fibrils from separate lamellae together. Bar = 0.2 μm. (D) Very high magnification (185,000×) quick-freeze, deep-etched electron micrograph of intralamellar collagen fibrils (long arrows) with beaded filaments (thick arrows) crisscrossing between fibrils and projecting three finger-like structures, which both appear to function in joining neighboring fibrils together. Bar = 0.1 μm. (C and D are from Hirsch M, et al. Three-dimensional supramolecular organization of the extracellular matrix in human and rabbit corneal stroma, as revealed by ultrarapid-freezing and deep-etching methods. Exp Eye Res 72:123, 2001.) |
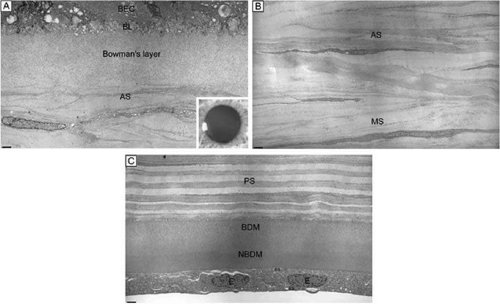 Figure 12. (A) The acellular Bowman’s layer and anterior-most portion of the cellular corneal stroma. Notice that the interweaving lamellae of cellular stroma branch and insert into the posterior surface of the Bowman’s layer forming an anterior corneal mosaic pattern after applying digital pressure on the corneal surface through the eyelid and fluorescein is instilled (inset). (B) The anterior third of the cellular corneal stroma showing the predominantly oblique lamellar orientation and the extensive vertical lamellar branches and interweaving. (C) The posterior third of the cellular corneal stroma, Descemet’s membrane and endothelium. Notice the parallel oriented and predominantly orthogonal arrangement of lamellae in this portion of corneal stroma. Although some collagen fibrils randomly insert into Descemet’s membrane, no specific pattern can be induced in this region of the cornea. BEC, basal epithelial cells; BL, basal lamina; AS, anterior stroma; MS, midstroma; PS, posterior stroma; BDM, banded portion of Descemet’s membrane; NBDM, non-banded portion of Descemet’s membrane; E, endothelial cells; ES, extracellular space. Bars = 1 μm. (TEM 4,750×.) |
 Figure 13. (A) The acellular Bowman’s layer in cross-section and frontal section (inset). Notice the random directionality of the 22-nm diameter collagen fibrils in this portion of the stroma. (B) The anterior third of the cellular corneal stroma in cross-section and frontal section (inset). Notice the thin and more obliquely oriented lamellae in this region of the corneal stroma and its lattice-like arrangement of 25-nm diameter collagen fibrils. (C) The posterior third of the cellular corneal stroma in cross-section and frontal section (inset). Notice the thick and parallel oriented lamellae in this region of the cornea stroma and its lattice-like arrangement of 25-nm diameter collagen fibrils. Also, note the orthogonal nature of lamellae (inset) compared to that in the inset from (B). Bars = 1 μm. (TEM 72,500×.) |
Although difficult to count exactly, the central cornea has been found to have approximately 300 corneal lamellae, whereas the peripheral cornea has approximately 500.61 Although these lamellae are generally described as running parallel to the corneal surface in orthogonal directions to one another, regional difference exist in their size, directionality, and amount of interweaving. The anterior third of the cellular corneal stroma has thinner (0.2- to 1.2-μm thick), narrower (0.5- to 30-μm wide), and mostly obliquely oriented lamellae with extensive vertical and horizontal interweaving (Figs. 12B and 13B), whereas the posterior two-thirds has thicker (1- to 2.5-μm thick), wider (100- to 200-μm wide), and mostly parallel oriented lamellae with only slight horizontal interweaving (Figs. 12C and 13C).60
Additionally, only lamellae in the posterior two-thirds of the cellular cornea stroma run in the classic orthogonal directions, whereas all of the lamellae of the anterior third and a minority of lamellae in the posterior two-thirds run in nonorthogonal directions. Because the lamellae in the anterior-most layers of cellular stroma branch so extensively, it is not surprising that they also interweave with (i.e., are attached to) the Bowman’s layer in a polygonal fashion, creating a mosaic appearance that can be seen on the anterior corneal surface under certain circumstances (Fig. 12A, inset). Finally, all corneal lamellae appear to stretch across the cornea from limbus to limbus where they turn and form an annulus approximately 1.5- to 2.0-mm wide around the cornea. This maintains the curvature of the cornea, while blending with limbal collagen fibrils.61,62,63
Corneal proteoglycans previously were referred to as extrafibrillar amorphous ground substance because their water soluble state made them difficult to fully delineate with light and electron microscopy (Fig. 14A). It was not until an electron-dense, cationic dye called cupromeronic blue and a critical-electrolyte-concentration of 0.1 M MgCl2 were used in combination to stain specifically for the sulfate-ester groups on corneal stromal proteoglycans that the shape, size, arrangement, and location of this material was observed with light and electron microscopy (Fig. 14B).64 Since that time, it has become quite apparent that corneal proteoglycans are not amorphous, but rather are tadpole-shaped structural molecules composed of a globular core protein (10 to 15 nm in diameter) with a covalently attached GAG sidechain or tail (7 nm wide × 45 to 70 nm in length). They are arranged in the cornea stroma orthogonal to collagen fibrils with a constant separation around 65 nm between each other along the fibrils. Their core proteins noncovalently bind to collagen fibrils in specific gap zones along the peripheral portions of the collagen fibril (core proteins with dermatan sulfate sidechains bind to d and e gap zones and those with keratan sulfate side chains bind to a and c gap zones).65 GAGs are highly negatively-charged, stiff polymers that extend into the interfibrillar space and form antiparallel duplexes with adjacent GAG sidechains, thereby crosslinking different collagen fibrils together by forming dumbbell-like structures (Fig. 14C).
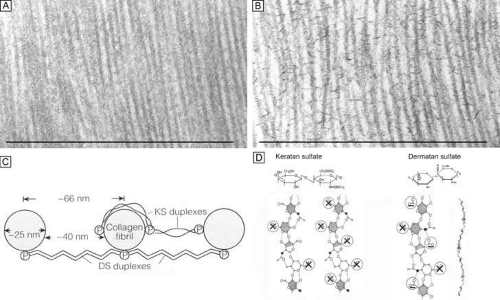 Figure 14. Frontal section TEMs (90,000×) of longitudinally running corneal stromal collagen fibrils in a lamellae without (A) and with (B) Cupromeronic blue staining. Notice the scattered “amorphous ground substance” in the interfibrillar spaces in (A), whereas in (B) duplexes of proteoglycans are clearly seen bridging collagen fibrils. Bars = 1 μm. (C) Diagram of how proteoglycans attach along the periphery of collagen fibrils via their core proteins (P) and how the GAGs duplex in an anti-parallel fashion in the interfibrillar space. (D) Diagram of the polymer backbones of keratan sulfate and dermatan sulfate. The top portion shows the primary structure of the repeating disaccharide units of keratan sulfate (top left) and dermatan sulfate (top right). The bottom portion of the diagram shows the secondary structures of each proteoglycan. Notice that in the human cornea ∼50% of keratan sulfate is in the normal-sulfated state (bottom, farthest left), whereas the other ∼50% is in the oversulfated state (bottom, center left). Also, notice that all three displayed proteoglycan polymers have similar backbones and therefore form similar secondary structures of a twofold helix (bottom, farthest right). Anionic charges = sulfate esters (X) and carboxylic acid (-). Bars = 1 μm. (C and D are modified from Scott JE. Proteoglycan: collagen interactions and corneal ultrastructure. Biochem Soc Trans 19:877, 1991.) |
Because the genes that produce the core proteins have now been cloned, four types of corneal proteoglycan core proteins have been identified: decorin, lumican, keratocan, and mimecan.66 Subsequently, decorin was found to contain a single dermatan sulfate GAG sidechain, whereas lumican and mimecan had single keratan sulfate GAG sidechains and keratocan had three keratan sulfate GAG sidechains. Thus, there are four known types of proteoglycan core proteins and only two types of GAGs: keratan sulfate (60%) and dermatan sulfate (40%), found in the human cornea stroma. The GAGs are polymers of repeating disaccharide units of galactose and N-acetylglucosamine or iduronic acid and N-acetylgalactosamine, respectively (Fig. 14D).65 Because GAG sidechains are posttranslationally added to the core protein portions in the golgi apparatus, there seems to be some flexibility in how long or how sulfated they can become depending of the type and functional needs of the connective tissue producing them. The human cornea is unique in that the GAGs are fibril-associated, small in length (keratan sulfate ≅45 nm and dermatan sulfate ≅70 nm), and with a higher proportion are oversulfated.
Interestingly, a comparative study of corneas from 12 mammalian species suggests that dermatan sulfate is the preferred proteoglycan in oxygen-rich environments such as thin corneas (e.g., mice) or in the anterior portion of thicker corneas (e.g., humans or rabbits), whereas keratin sulfate is a functional substitute produced through an alternate metabolic pathway in thicker corneas, especially in the posterior portion where oxygen levels may be dramatically reduced (Fig. 15, inset).48,66 Functionally, this duality is quite useful because dermatan sulfate appears to be more efficient at holding water as it absorbs less total amounts of water, but most in a tightly-bound state (i.e., in a nonfreezable state) than does keratin sulfate.47,67 These explanations are consistent with the fact that dermatan sulfate is highest in amount in the anterior portion of the corneal stroma in humans (region of highest oxygen tension and most affected by evaporation), whereas keratin sulfate is highest in amount in the posterior portion of the corneal stroma (region with the lowest oxygen tension, least affected by evaporation, and area where the need for loosely-bound water is required for transport across the endothelium through the endothelial cell metabolic pump) (Fig. 15).
 Figure 15. Diagram demonstrating the regional differences in the corneal stroma for the proportion of the two types of corneal proteoglycans and the water absorbing properties in these regions. (Inset) Diagram of the metabolic pathways for dermatan sulfate and keratan sulfate production. Notice that the supply of oxygen is the initial factor whether dermatan sulfate is made through an aerobic pathway or whether keratan sulfate is made through an anaerobic alternative pathway. (Modified from Scott JE. Oxygen and the connective tissues. TIBS 17:340, 1992.) |
The corneal stroma is maintained by a closed, highly-organized syncytium of keratocytes communicating with each other through gap junctions present on their long dendritic processes.68 Keratocytes occupy 10% to 40% of the stromal volume (decreases from 40% in infancy to 10% in adulthood) that on two-dimensional, cross-sectional views appear as flattened (cell body = 20 μm in length × 1 μm in height), quiescent (i.e., scant intracytoplasmic organelles) cells lying between corneal lamella (Fig. 16A, B). In actuality, keratocytes are three-dimensional, stellate-shaped cells composed of a cell body (≅1 × 15 × 20 μm) with numerous dendritic-processes that extend up to 50 μm in length from the cell body. Two-dimensional tangential sections of the normal cornea suggest that these cells are more highly metabolically active in the resting state than initially presumed; in tangential sections (cell body view = 15 μm width × 20 μm), an abundance of cytoplasmic organelles is commonly seen (Fig. 16C, D).69
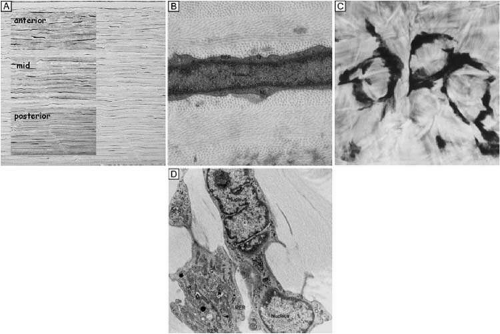 Figure 16. Light and TEM micrographs shown in cross and frontal-sectional views of the cellular corneal stroma demonstrating differences in the appearance of keratocytes depending on the cut, or perspective, of the section. (A) Cross-sectional light microscopy shows that keratocytes are primarily obliquely aligned to corneal surface in the anterior one-third of cellular corneal stroma and are aligned parallel to the corneal surface in the posterior two-thirds. (B) Cross-sectional TEM additionally shows that keratocyte nuclei occupy most of the area of the keratocyte seen in this perspective with only a thin rim of surrounding cytoplasm that contains only small numbers of cytoplasmic organelles. (C) From a tangential perspective, frontal-section light microscopy shows that keratocytes are arranged in a circular fashion. (D) Frontal-section TEM additionally shows that supposedly quiescent keratacytes may be more active in the baseline state than initially thought as an extensive amount of cytoplasmic organelles can be seen in this view. M, mitochondria; RER, rough endoplasmic reticulum; V, vacuoles. *Main portion of nucleus that contains nucleolus. (Modified from Muller LJ, et al. Novel aspects of the ultrastructural organization of human corneal keratocytes. Invest Ophthalmol Vis Sci 36:2557, 1995.) |
Tangential sections also have shown more clearly that the anterior stromal keratocytes contain twice the number of mitochondria than the posterior two-thirds of the stroma, which correlates with the oxygen tension levels in the cornea. It also has demonstrated that a higher density or volume of cells reside in the anterior stroma than in the mid or posterior stroma (Fig. 17). Moreover, these views make it easy to appreciate that, in all levels, the keratocytes are highly spatially-ordered as they turn in a clockwise direction like a corkscrew.
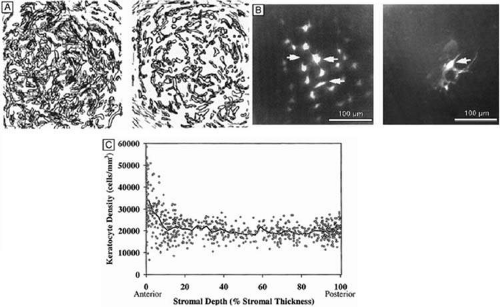 Figure 17. (A) Reconstruction of keratocyte outlines seen in frontal-section in the anterior and posterior thirds of the cellular corneal stroma. (Modified from Muller LJ, et al. Novel aspects of the ultrastructural organization of human corneal keratocytes. Invest Ophthalmol Vis Sci 36:2557, 1995.) (B) Fluorescent-dye spreading between many adjacent keratocytes in rabbit (center left) and human corneas (center right), which demonstrates the intimate importance of gap junctions in how keratocytes communicate with one another. (From Watsky MA. Keratocyte gap junctional dye spread in normal and wounded rabbit corneas and human corneas. Invest Ophthalmol Vis Sci 36:S22, 1995.) (C) Mean keratocyte density along the depth of central cellular corneal stroma. Notice the zone of increased density the closer one gets to the epithelial-stromal junction. Perhaps this is due to baseline, normal epithelial-stromal interactions. (From Patel SV, et al. Normal human keratocyte density and corneal thickness measurement by using confocal microscopy in vivo. Invest Ophthalmol Vis Sci 42:333, 2001.) |
In vivo confocal microscopy of normal human corneas has shown keratocyte densities average around 20,000 keratocytes/mm3 (or an area of 328 cells/mm2). The focal zone of increased cell density directly under the epithelial surface averages 35,000 keratocytes/mm3 in the anterior-most 9-μm layer and gradually tapers to 20,000 keratocytes/mm3 over the initial 60 μm in depth (Fig. 17C).70 Confocal microscopy also concurs with previous studies as it shows that stromal cellular density decreases with age at approximately 0.45% per year of life. Interestingly, studies using immunohistochemistry on animal corneas71,72 and ultrastructural studies on human corneas69,72 suggest that not all the cells in the corneal stroma are keratocytes (i.e., fibrocytes), but some are one of three types of bone marrow-derived immune cells (“professional” dendritic cells, “nonprofessional” dendritic cells, and histiocytes) (Fig. 18). Thus, although the majority of cells in the stroma are keratocytes, which maintain the corneal stroma by synthesizing and secreting collagen type I, III, V, VI, XII, XIV, keratan sulfate, dermatan sulfate, and matrix metalloproteinases, a minority of cells in the stroma are also “professional” stromal dendritic cells, “nonprofessional” stromal dendritic cells, or stromal histiocytes. They appear to play a pivotal role in the induction of immune tolerance versus immune initiation into cell-mediated immune pathways. Additionally, the stromal histiocytes play a role in innate immunity as phagocytic effectors cells.
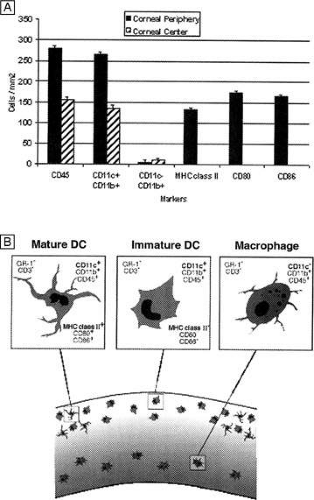 Figure 18. The mouse cornea suggests that 5% to 10% of the cells in the cellular corneal stroma are actually one of three types of bone marrow-derived immune cells. The anterior one-third of the cellular corneal stroma contains professional (MHC-positive) dendritic antigen-presenting cells in periphery of the cornea, whereas nonprofessional (MHC-negative) dendritic cells are present in both the periphery and center of the anterior cornea. The posterior-most regions of the cellular corneal stroma appear to contain macrophages in the periphery and center of the cornea. (From Hamrah P, et al. The corneal stroma is endowed with a significant number of resident dendritic cells. Invest Ophthalmol Vis Sci 44:581, 2003.) |
Two ion channels similar to those found in excitable cells have been characterized in keratocytes isolated from corneal stroma.73 These channels include a delayed rectifer K+ channel and a tetrodotoxin-sensitive Na+ channel. The physiologic function of these channels has not been determined, but they may function in maintaining the ionic balance of the cells in their inactive state. In addition, because these channels allow the keratocytes to elicit action potentials when injected with a clamping current, it is possible they may act as intercellular signalers via the gap junctions between cells. This could be a route for information exchange between cells in different regions of the cornea.
Corneal Edema
The Donnan effect states that the swelling pressure in a charged gel, like the corneal stroma, results from ionic imbalances. The fixed negative charges on corneal proteoglycan GAG sidechains (one carboxylic acid and one sulfate ester sidechain per disaccharide repeat on a dermatan sulfate GAG polymer, and one or two sulfate ester sidechains per disaccharide repeat on a keratin sulfate GAG polymer) have a central role in this effect as the antiparallel GAG duplexes (tertiary structure) produce long-range electrostatic repulsive forces that induce an expansive force termed swelling pressure (SP). Because the corneal stroma also has a cohesive tensile strength that resists this expansion, the normal SP of the nonedematous corneal stroma is around 55 mm Hg.74,75 If the stroma is further compressed (e.g., increasing intraocular pressure [IOP] or mechanical applanation) or expanded (e.g., corneal edema), the SP will correspondingly increase or decrease, respectively.
Conversely, the negatively-charged GAG sidechains also form a twofold helix in aqueous solution (secondary structure) that attracts and binds Na+ counterions, which results in an osmotic effect leading to the diffusion and subsequent absorption of water to proteoglycans via the Na+ cations. Thus, the central corneal thickness is maintained around its average value of 520 μm because the fixed negatively-charged proteoglycans induce a swelling pressure through anionic repulsive forces. Additionally, the hydration level of corneal stroma is maintained around 78% water because these proteoglycans also imbibe water through cationic attractive forces.45
Under normal circumstances, the negative pressure drawing fluid into the cornea, called the imbibition pressure (IP) of the corneal stroma, is around -40 mm Hg.76 This implies that the negative charges on corneal proteoglycans are only about one-quarter (≅27 %) saturated, or bound, with water with the remaining unbound proportion wanting to bind more Na+ and absorb more water if given the opportunity. Fortunately, the highly impermeable epithelium and mildly impermeable endothelium keep the diffusion of electrolytes and fluid flow in the stroma to such a low level [resistance to diffusion of electrolytes and fluid flow = epithelium (2000) >> endothelium (10) > stroma (1)] that the continuously working endothelial cell metabolic pump can maintain stromal hydration in the normal range. Although IP = SP when corneas are in the ex vivo state, IP actually is lower than SP in the in vivo state because the hydrostatic pressure induced by the IOP must now be accounted for. This is best represented by the equation IP = IOP – SP76 and explains why the hydration level of patient’s cornea is not only dependent on having normal barrier functions, but also on having a normal IOP. Therefore, a loss of corneal barrier function, an IOP ≥55 mm Hg, or a combination of the two results in corneal edema (Fig. 19).77
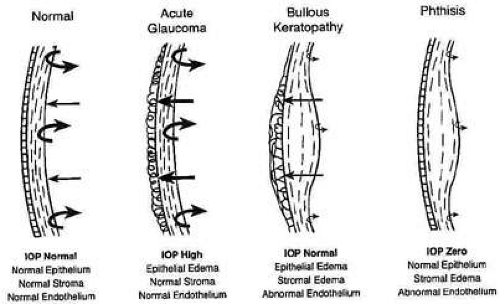 Figure 19. Diagram demonstrating the delicate balance between stromal swelling pressure, endothelial pump function, and intraocular pressure. Usually if endothelial cell pump function fails and IOP remains normal, slightly decreases, or increases, both stromal and epithelial edema occurs (center right). Only when IOP increases above the swelling pressure of the stroma and the endothelium functions normal do we see epithelial cell edema alone (center left) and only when IOP is zero and the endothelium functions abnormally do we see stromal edema alone (far right). (From Hatton MP, et al. Corneal edema in ocular hypotony. Exp Eye Res 78:549, 2004.) |
Corneal edema is a term often used loosely and nonspecifically by clinicians, but literally refers to a cornea that is more hydrated than normal (i.e., >78% water). The topic of corneal edema is important for clinicians to understand because it affects the architecture and function of the epithelium and the stroma. Epithelial edema clinically causes a hazy microcystic appearance to occur in the epithelium in mild to moderate cases, significantly decreasing vision and increasing glare. It also can cause the development of large, painful, subepithelial bullae in severe cases. These changes correlate histopathologically with hydropic basal epithelial cell degenerative changes and the development of extraepithelial cellular fluid filled spaces (e.g., cysts and bullae). Interestingly, if bullae are chronically present, a fibrocollagenous degenerative pannus often times will grow into the subepithelial space, decreasing vision further while reducing the pain.
Stromal edema clinically appears as a painless, hazy thickening of the corneal stroma resulting in a mild to moderate reduction in visual acuity and an increase in glare. At the same time, Descemet’s membrane folds commonly appear on the posterior surface of the cornea. Histopathologically, these changes correlate with the light microscopic findings of thickening of the corneal stroma in the posterior cross-sectional direction with loss of artifacteous stromal clefting.78 Ultrastructural and biochemical studies have further shown that stromal edema causes an increase in the distance and disruption of spatial order between collagen fibrils,78 a decrease in the refractive index of the extracellular matrix,79 hydropic degenerative changes or cell lysis in the resident keratocyte population,80 and a loss of proteoglycans.81
Although different proportions of the two types of negatively-charged proteoglycan may account for the higher hydration levels in the posterior stroma compared to the anterior stroma,46,82 it appears that the directional orientation of the collagen fibrils probably has the greatest influence on how much each region thickens, or swells, as a result of increased hydration levels. Because the collagenous architecture of the stroma (i.e., limbus-to-limbus directional orientation of collagen fibrils) highly resists circumferential expansion, only anterior-posterior expansion occurs in the human cornea, mostly in the posterior direction. This latter fact occurs because extensive lamellar interweaving occurs in the anterior portions of the corneal stroma, whereas weak bridging filaments (i.e., type VI and FACIT collagens) occur diffusely throughout the entire corneal stroma. Furthermore, this lamellar interweaving also explains why the anterior third of the cornea mildly swells and actually maintains its anterior corneal curvature even when the remaining stroma swells to up three times its normal thickness.80 Because fibrotic corneal scars have random directionally-oriented interweaving collagen fibrils, they also have been found to resist swelling under edematous conditions.83
Therefore, although it is commonly stated that corneal thickness and interfibrillar spacing increase in a linear fashion to the hydration level of the corneal stroma,75,78 one needs to be aware that this relationship mainly applies to the mid and posterior stromal regions.
Finally, although both epithelial and stromal edema commonly co-exist together, there are two notable exceptions that are important to understand. Because the epithelium lacks fixed negatively-charged proteoglycans and has a different set of cohesive mechanical strengths than the stroma (e.g., intercellular desmosomal junctions), its state of hydration is mainly dictated by IOP levels.84 Conversely, because collagen fibrils in corneal stroma are anchored at the limbus for 360 degrees, they exert increasing cohesive tensile mechanical forces on the corneal stroma (i.e., compression of stromal tissue) as the IOP elevates above normal. This results in the transmission of edema to the epithelial surface in cases of high IOP. Therefore, if IOP is ≥55 mm Hg with normal endothelial barrier and pump function, epithelial edema usually occurs by itself. In contrast, if endothelial cell dysfunction and hypotony (IOP ∼0 mm Hg) occur together, then stromal edema occurs alone (see Fig. 19).
MICROSCOPIC ANATOMY, ULTRASTRUCTURE, AND PHYSIOLOGY OF THE ENDOTHELIUM
The endothelium of the infant cornea is composed of a single layer of approximately 500,000 neural crest-derived cells, each measuring around 5 μm in thickness by 20 μm in diameter and cover a surface area of 250 μm2.58,85 The cells lie on the posterior surface of the cornea and form an irregular polygonal mosaic. The tangential appearance of each corneal endothelial cell is uniquely irregular, usually uniform in size to one another, and typically six-sided (i.e., hexagons). They abut one another in an interdigitating fashion with 20 nm wide intercellular space between each other. The intercellular space is known to contain discontinuous apical tight junctions (macula occludens) and lateral gap junctions, thereby forming an incomplete barrier to diffusion of small molecules. As corneal endothelial cells have numerous cytoplasmic organelles, particularly mitochondrial organelles, they have been inferred to have the second highest aerobic metabolic rate of cells in the eye next to retinal photoreceptors.58
At birth, the central endothelial cell density of the cornea is around 5,000 cells/mm2.85 Because the corneal endothelium has very limited regenerative capabilities, there is a well-documented decline in central endothelial cell density with age that typical involves two phases: a rapid and a slow component.85,86 Due to corneal growth and age-related, or developmental, selective cell death, during the fast component, the central endothelial cell density decreases exponentially to about 3,500 cells/mm2 by age 5 years and 3,000 cells/mm2 by age 20 years.86 Thereafter, a slow component occurs where central endothelial cell density decreases to a linear steady rate of 0.6% per year, resulting in cell counts around 2,500 cell/mm2 in senescence (Fig. 20).85.86 Because endothelium maintains its continuity by migration and expansion of surviving cells, it is not surprising that the percentage of hexagonal cells decreases (pleomorphism) and the coefficient of variation of cell area increases (polymegathism) with age.86
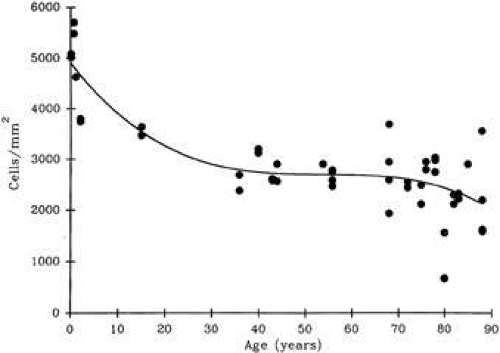 Figure 20. Scatterplot with best fit curve showing the average central corneal endothelial cell density for normal, healthy eyes of different ages. (From Williams KK, et al. Correlation of histologic corneal endothelial cells counts with specular microscopic cell density. Arch Ophthalmol 110:1146, 1992.) |
It is important when reviewing this information to realize that these are average central corneal endothelial cell measurements from predominantly Caucasian U.S. populations, as racial and geographic differences can exist. Also, understand that this data applies only to central corneal endothelial measurements because recent work has shown that higher cell densities can typically be found in more peripheral aspects of the cornea (Fig. 21).87 Therefore, based on these studies, it appears that central corneal endothelial cell numbers decrease on average about 50% from birth to death in normal subjects. Because corneal decompensation typically does not occur until central values reach around 500 cells/mm2, there appears to be plenty of cellular reserve potential remaining after an average human life span.86 Estimates suggest that healthy, normal human corneal endothelium could maintain corneal clarity up to a minimum of 215 years of life, if humans lived that long.85
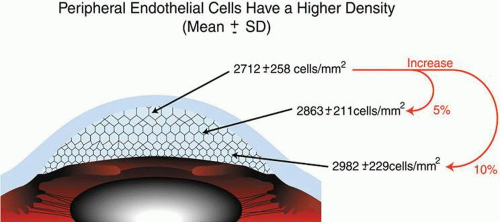 Figure 21. Diagram illustrating the central, paracentral, and peripheral corneal endothelial cell densities in healthy, normal subjects (From Amann J, et al. Increased endothelial cell density in the paracentral and peripheral regions of the human cornea. Am J Ophthalmol 135:584, 2003.)
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
 Get Clinical Tree app for offline access
Get Clinical Tree app for offline access

|