Conjunctiva
Brian. J. Song
Nahyoung Grace Lee
Samir M. Haq
James D. Zieske
Stefan D. Trocme
The conjunctiva is a thin, highly vascularized mucous membrane that extends from the margins of the upper and lower eyelids to the limbus, a transition zone at the border of the cornea on the anterior surface of the eye. Its name is derived from the fact that it serves as an attachment between the eyelids and the globe.1 Together, the conjunctiva, limbus, and the cornea comprise the ocular surface.2 Histologically, the conjunctiva is composed of two layers: (a) the conjunctival epithelium and (b) the substantia propria (Fig. 29.1). The conjunctival epithelium consists of nonkeratinized, stratified cells that act as a protective barrier on the ocular surface. Deep to the conjunctival epithelium, the substantia propria provides structural support for the overlying epithelium and houses the immune cells of the ocular surface within its lymphoid tissue.
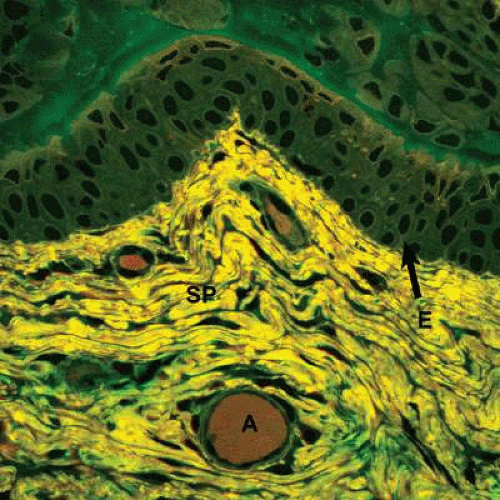 Figure 29.1. High-power view of the conjunctiva showing the stratified cells of the epithelium (E) in green overlying the connective tissue of the substantia propria (SP) in yellow. A conjunctival artery (A) is readily visible within the connective tissue (Confocal H&E ×800). (Courtesy of Dr. Donald W. Pottle, Schepens Eye Research Institute.) |
Functionally, the conjunctiva plays an important role in both ocular immunology and in the maintenance of the adjacent cornea. Four main reasons account for the remarkable ability of the conjunctiva to combat infections: (a) it is highly vascular; (b) it contains different cell types that participate in defensive inflammatory reactions; (c) it has many immunocompetent cells that act as a source of immunoglobulins; and (d) the surface anatomy (microvilli) and biochemistry (enzymatic activity) of the conjunctival cells enable it to engulf and neutralize viruses and other foreign particles.3,4 In addition, the conjunctiva supplies epithelial cells and blood vessels to promote healing, whereas its ability to regulate mucin and fluid secretion helps stabilize the tear film and maintain corneal transparency.5,6
Clinically, the conjunctiva possesses many features that aid both the surgical and medical management of patients. The loosely adherent properties of the bulbar surface allow the conjunctiva to be used to great advantage in glaucoma surgery. Conjunctival autografts can be carried out from one eye to the other to repair leaking filtration blebs and to hasten epithelial recovery after chemical burns have damaged the corneal surface.7 A conjunctival flap placed over a nonhealing or infected cornea can help promote healing.8 The vascularity of the conjunctiva also promotes rapid healing, which ensures the success of many surgical procedures. Another exceptional advantage of the conjunctiva is the ease with which conjunctival biopsy material can be obtained, causing little discomfort to the patient and producing virtually no loss of tissue integrity. The conjunctiva also can serve as a marker for underlying systemic diseases. For example, a pathognomonic conjunctival vasculopathy can be seen in patients with sickle cell anemia, whereas a distinctive bluish-black pigmentation is indicative of ochronosis (Fig. 29.2).9,10 Crystalline deposits in the conjunctiva may be a sign of cystinosis, and even nutrient deficiencies, such as hypovitaminosis A, can be diagnosed through the presence of Bitot spots on the ocular surface.11,12 The transparent nature of the conjunctiva can even allow the astute clinician to diagnose rare infections, such as Loa loa filariasis, by identifying the worm crawling under the conjunctival surface.13
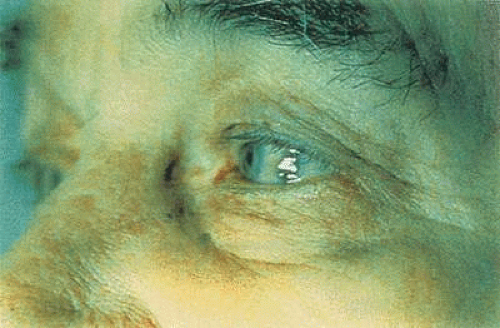 Figure 29.2. Bluish-black pigmentation characteristic of ocular ochronosis seen here on the lateral side of the bulbar region. (Reproduced with permission from Gold DH, Weingeist TA(eds): Color Atlas of the Eye in Systemic Disease. Philadelphia: Lippincott Williams & Wilkins, 2001:313.) |
In this chapter, we will review conjunctival gross anatomy, histology, pathophysiologic responses to inflammation and wound healing, and changes of the conjunctiva associated with aging.
Gross Anatomy
The conjunctiva lines the posterior surface of the upper and lower eyelids and covers the anterior portion of the sclera. From the inner surface of the lid, it is reflected onto the globe above and below, forming two recesses: the superior and inferior fornix. This reflection of the conjunctiva from the lids to the globe also creates a potential space called the conjunctival sac, which opens out through the palpebral fissure between the upper and lower eyelids. The superior fornix is located at the level of the orbital margin, 8 to 10 mm above the limbus (Fig. 29.3A). The inferior fornix is approximately 8 mm below the limbus (Fig. 29.3B). Medially, the forniceal structures are replaced by specialized areas known as the caruncle and the plica semilunaris (Fig. 29.3C). The absence of the fornix on the medial side is necessary to allow the inferior punctum to dip and drain from the superficial tear fluid layer.1 Laterally, the fornices are quite deep, extending behind the equator of the globe approximately 14 mm from the limbus (Fig. 29.3D).14
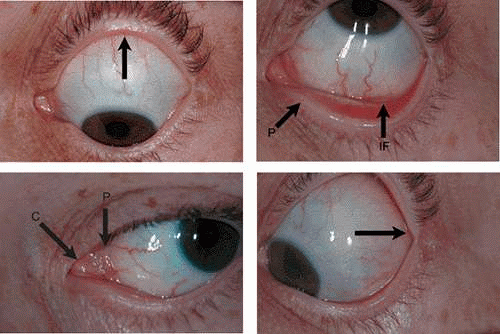 Figure 29.3. A. Region of the superior fornix (arrow) located approximately 8 to 10 mm from the limbus. B. Region of the inferior fornix (IF) located approximately 8 mm from the limbus. The opening of the punctum (P) is shown on the medial side of the marginal conjunctiva of the lower lid. C. Medial region of the eye showing the two specialized areas of conjunctiva: the lacrimal caruncle (C) and the plica semilunaris (P).D. Region of the lateral fornix (arrow). The lateral fornix is quite deep, and the actual recess is not readily visible. |
Anatomically, the conjunctiva is divided into three regions: (a) palpebral; (b) forniceal; and (c) bulbar (Fig. 29.4). The palpebral conjunctiva begins at the mucocutaneous junction, located at the posterior end of the eyelid margin where the epidermis of the eyelid abruptly transforms into the conjunctiva proper. From there, it lines the inner surface of the eyelids, where it adheres tightly to the posterior surface of the tarsal plates before continuing on toward the fornices. The palpebral conjunctiva also can be subdivided further into marginal, tarsal, and orbital zones.1 The marginal conjunctiva, as its name suggests, begins at the mucocutaneous junction of the skin and the conjunctiva proper. The punctum opens into the medial side of the marginal conjunctiva of the lower lid, creating a communication between the conjunctival sac and the inferior nasal meatus via the lacrimal system. The tarsal conjunctiva is the portion of the palpebral conjunctiva that is affixed to the tarsal plates. It is highly vascular and, because it is translucent, the tarsal glands, which are embedded in the tarsal plates, can be seen through it as yellowish streaks (Fig. 29.5A and B). Although the tarsal conjunctiva is firmly attached to the entire superior tarsus, it is only attached to half the width of the inferior tarsus. The orbital conjunctiva is the remaining region of the palpebral conjunctiva, extending from the peripheral edge of the tarsus to the fornix. Movement of the eye causes a horizontal folding of the orbital conjunctiva that is most pronounced when the eye is open.1 Functionally, the palpebral conjunctiva is an area in which reactive pathology may be seen. In particular, follicle and papilla formation occur in this region in conjunction with various forms of conjunctivitis.15,16,17,18
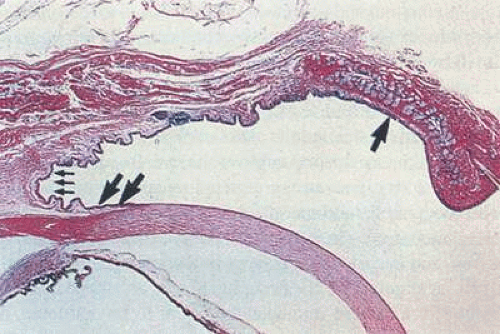 Figure 29.4. Sagittal section showing the three main divisions of the conjunctiva. The palpebral conjunctiva (single arrow) covers the posterior portion of the eyelid and transitions into the forniceal conjunctiva (triple arrow) in the recess where the conjunctiva reflects from the eyelid onto the surface of the globe. The bulbar conjunctiva (double arrow) extends over the anterior globe from the fornix to the limbus (H&E ×2.5). (Reproduced with permission from Sternberg SS(ed): Histology for Pathologists, 2nd ed. Philadelphia: Lippincott-Raven, 1997:321.) |
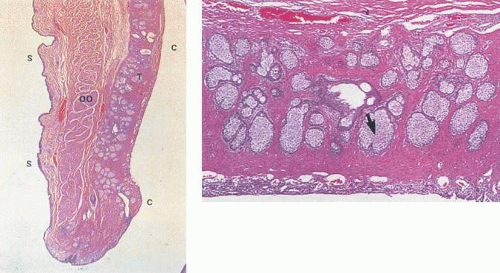 Figure 29.5. A. Sagittal section of the eyelid showing the skin (S) to the left and the palpebral conjunctiva (C) lining the posterior lid and tarsus (T) on the right. The tarsal (meibomian) glands can be seen within the tarsus. The orbicularis oculi (OO) muscle is clearly labeled. B. The meibomian glands (arrow) within the dense fibrous tissue of the tarsus are seen here. The tarsal conjunctiva lining the tarsus is seen at the bottom of this image (A. H&E ×3.3;B. H&E ×13.2). (Reproduced with permission from Sternberg SS, ed. Histology for pathologists, 2nd ed. Philadelphia: Lippincott-Raven, 1997:321,334.) |
The forniceal conjunctiva is located in the superior and inferior forniceal recesses between the palpebral and bulbar conjunctiva. Unlike the palpebral conjunctiva, the forniceal conjunctiva is loosely attached to the underlying tissue, specifically the fascial sheaths of the levator palpebrae superioris and rectus muscles. This loose attachment allows the forniceal conjunctiva to move with the globe and the eyelids when these muscles are contracted.14 The lateral side of the superior forniceal conjunctiva is the site at which the lacrimal gland duct opens.
The bulbar conjunctiva extends from the forniceal recesses to the limbus. It is so thin and translucent that the underlying sclera can be seen through it. The bulbar conjunctiva is loosely attached to the sclera by areolar tissue, which allows the eye to move freely in all directions. It is also attached to the tendons of the rectus muscles, which in turn are covered by Tenon capsule, a dense, collagenous fascia-like layer that extends over the anterior globe from the insertions of the rectus muscles.19 Approximately 3 mm from the limbus, the bulbar conjunctiva fuses with Tenon capsule and the sclera, making it difficult to grasp the conjunctiva in the paralimbal region.20 This attachment is routinely encountered during the dissection of a limbal-based conjunctival flap in ocular surgery. Table 29.1 summarizes the anatomic features of each region of the conjunctiva.
TABLE 29.1. Anatomic divisions of the conjunctiva | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Arteries
The conjunctival arteries originate from two main sources: (a) the palpebral branches of the ophthalmic and lacrimal arteries; and (b) the anterior ciliary artery. Both the lacrimal and the anterior ciliary arteries originate from the ophthalmic artery, which is derived from the internal carotid artery.1 The superior and inferior medial palpebral arteries branch off the ophthalmic artery on the nasal side of the globe, whereas the superior and inferior lateral palpebral arteries stem from the lacrimal arteries on the temporal side. The corresponding lateral and medial palpebral arteries then join within the eyelid to form the peripheral and marginal palpebral (tarsal) arcades (Fig. 29.6). Together, these two palpebral arcades supply the entire conjunctiva, with the exception of the paralimbal region. As its name implies, the marginal arcade runs along the marginal border of the tarsus, more proximal to the lid margin than the peripheral arcade. The marginal arcade is the larger of the two palpebral arcades. It is located approximately 3 mm from the mucocutaneous junction of the eyelid and runs between the tarsus and the orbicularis oculi muscle.14 The perforating arteries from the marginal palpebral arcade pass through the tarsus, reaching the subconjunctival space in the region of the subtarsal groove to form the marginal and tarsal vessels. On the posterior tarsus of the upper lid, the vessels from the marginal arcade ascend to join the peripheral vessels (from the peripheral arcade) and form a post-tarsal plexus that supplies the palpebral conjunctiva.
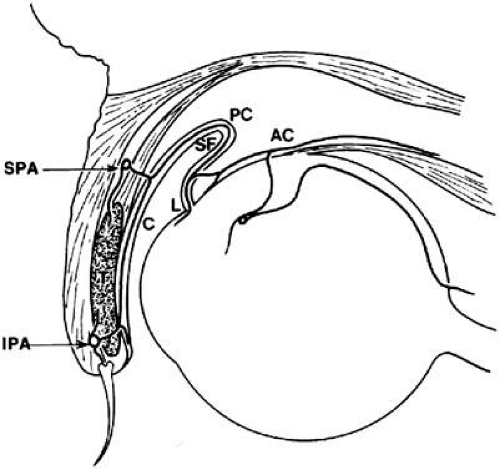 Figure 29.6. Arterial supply of the conjunctiva. Marginal (MA) and peripheral (PA) arcades run across the inferior and superior borders of the tarsus (T) respectively. The posterior conjunctival artery (PC) travels from the palpebral conjunctiva (C), over the superior fornix (SF), and toward the limbus (L). The anterior ciliary artery (AC) supplies the paralimbal region. (Modified with permission from Duke-Elder S. System of ophthalmology, Vol. 2. St. Louis: C.V. Mosby, 1961:547.) |
The peripheral palpebral arcade runs along the peripheral border of the tarsus within the eyelid. Branches of the peripheral arcade are responsible for most of the blood supply to the forniceal conjunctiva. The perforating vessels from the peripheral palpebral arcade pierce the Müller muscle and divide into descending and ascending branches.1 The descending branches supply the proximal two-thirds of the tarsal conjunctiva and also join with the vessels from the marginal arcade, as mentioned previously. As a result, the rich blood supply to the tarsal conjunctiva gives it a characteristic red color, compared to the relatively colorless bulbar conjunctiva. The ascending branches pass around the fornices to become the posterior conjunctival arteries. These vessels anastomose with the anterior conjunctival arteries approximately 4 mm from the limbus, to supply most of the bulbar conjunctiva except for the paralimbal region.1 The peripheral arcade of the lower lid is absent in some people. In these cases, the marginal arcade or the anterior ciliary arteries supply the inferior tarsal, forniceal, and bulbar conjunctiva.
The anterior ciliary arteries provide the second major source of blood to the conjunctiva. The branches of the anterior ciliary artery supply the paralimbal region of the bulbar conjunctiva. These arteries originate from the muscular branches of the ophthalmic artery, travel along the tendon of the rectus muscles, and through the Tenon capsule to give off the anterior conjunctival arteries just before piercing the globe. At this point, the anterior ciliary arteries send episcleral branches to a pericorneal plexus and to neighboring regions of the bulbar conjunctiva in the limbal area. Free anastomosis occurs in this area between the anterior conjunctival vessels and the terminal branches of the posterior conjunctival vessels. Thus, the superficial and deep systems in the limbal area are closely connected. Clinically, this is an area of diagnostic importance. The superficial posterior vessels become engorged during inflammation or infection to give the bulbar conjunctiva a bright red appearance that fades from the fornix toward the limbus.1 In keratitis or iritis, the deeper ciliary vessels are hyperemic, which leads to a characteristic circumcorneal injection that blanches with pressure.1 Figure 29.7 shows a high-power view of a conjunctival artery within the substantia propria.
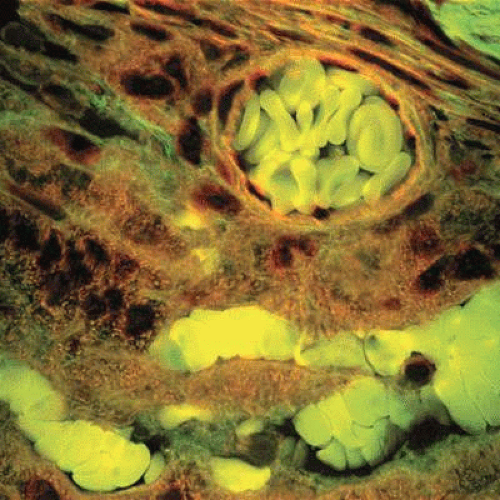 Figure 29.7. High-power view of a conjunctival artery in the substantia propria imaged here using three confocal channels. The biconcave shape of the red blood cells (light green) can be appreciated within the lumen of the vessel (Confocal H&E ×1,600). (Courtesy of Dr. Donald W. Pottle, Schepens Eye Research Institute.) |
Veins
The veins of the conjunctiva are more numerous than are the arteries.1 The conjunctival veins accompany their corresponding arteries and ultimately drain into either the palpebral veins or the superior and inferior ophthalmic veins. The major portion of the drainage from the tarsal, forniceal, and bulbar conjunctiva is directed toward the palpebral veins. Some of the tarsal veins empty independently into the superior and inferior ophthalmic veins. Outflow from the circum-corneal region goes to those veins that serve the extraocular muscles.
Small blood vessels of the bulbar conjunctiva have arteriovenous communications.20 The communicating vessels may be tortuous and uneven in caliber, but they are usually larger in diameter than are capillaries. Each gives off capillary branches proximally and receives capillaries distally. They are not true arteriovenous anastomoses, because they do not possess muscular walls that would render them capable of responding to chemical agents. Occasionally, the conjunctival blood vessels may become damaged, causing a spontaneous subconjunctival hemorrhage. These hemorrhages rarely have any association with systemic diseases and usually absorb in a matter of 1 or 2 weeks. On electron microscopy, the majority of the conjunctival capillaries have a thick, continuous wall with few fenestrations (Fig. 29.8).
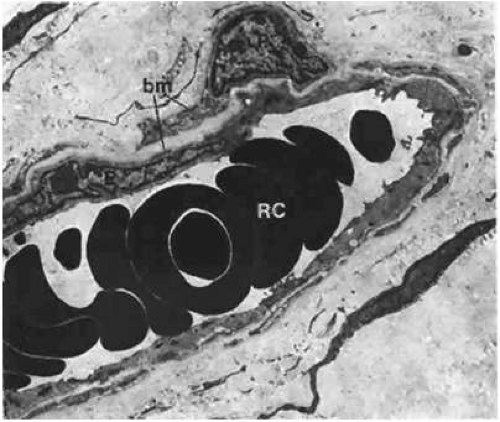 Figure 29.8. A blood capillary in the stroma of the bulbar conjunctiva. This is a nonfenestrated type, but a fenestrated type also is present in the conjunctival stroma. Both endothelium (E) and pericytes (P) are surrounded by a basement membrane (bm), and the lumen contains erythrocytes (RC) (×6,300). |
Aqueous Veins
The aqueous veins, as first described by Ascher, are vessel-like structures connecting the canal of Schlemm and its outlets with the conjunctival and episcleral vessels of the deep scleral meshwork.21,22 These veins vary in diameter from 0.01 to 0.1 mm and are easily identifiable on close biomicroscopic surface examination of the conjunctiva. They usually are found near the nasal limbus and appear hook-shaped when they first come off the sclera. The aqueous veins transport a clear fluid as they travel a short course of approximately 1 cm from the paralimbal region toward the equator of the bulbus. The blood of the episcleral veins may become diluted when joining the aqueous veins; alternatively, the clear fluid and blood may run side by side unmixed, forming a laminated vein. The aqueous veins serve as exit channels for the aqueous humor, and thus, they have a role in the pathophysiology of glaucoma.21,22 Histochemically, the expression of alkaline phosphatase is much weaker in the endothelium of the aqueous veins when compared to normal blood vessels.23 As a result, disparities in alkaline phosphatase activity may be helpful in distinguishing aqueous veins from other vessels of the ocular surface.
Lymphatics
The vast lymphatic network of the conjunctiva is arranged in two plexuses: (a) a superficial plexus consisting of small vessels below the capillaries; and (b) a deeper plexus of larger vessels in the fibrous portion of the substantia propria.20 These vessels are important in the mediation of the immunologic reactions that occur in certain ocular diseases and surgical conditions.
The superficial plexus receives lymphatic drainage from the limbal area. It has larger collector channels that run circumferentially 7 to 8 mm behind the limbus, forming an incomplete pericorneal lymphatic ring. Its other lymphatic drainage channels include a recurrent nasal group that drains the upper nasal quadrant and a descending temporal group. Both groups drain by way of the medial canthus. A large collecting vessel from the inferior fornix empties by way of the lateral canthus. In general, laterally placed lymph vessels flow to the preauricular and intraparotid lymph nodes, whereas medially placed vessels flow to the submaxillary lymph nodes. Lymph vessels may be visible on biomicroscopic examination in certain conditions, such as scleredema adultorum, and some may even fill with blood to give the appearance of dilated veins.24,25 Electron microscopy reveals that these lymphatics differ from blood capillaries structurally. The endothelium is very thin but is not fenestrated, and pericytes are usually absent. In addition, their intercellular junctions are less well formed than in capillaries, and the basement membrane tends to be interrupted or absent (Fig. 29.9).
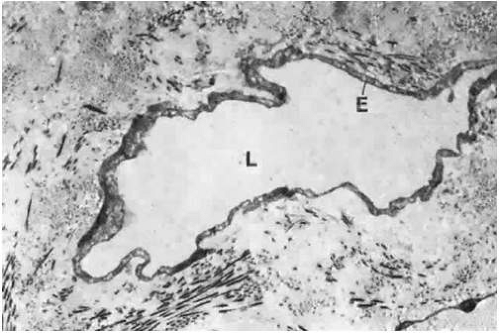 Figure 29.9. A lymphatic capillary in the substantia propria of the bulbar conjunctiva. The endothelium (E) is thin, and the basement membrane is often not well developed (as in this picture). The lumen (L) usually contains no erythrocytes (×13,700). |
Nerve Supply
The nerves that supply the eyelid also innervate the conjunctiva. These nerves include the infratrochlear branch of the nasociliary nerve, the lacrimal nerve, the supratrochlear and supraorbital branches of the frontal nerve, and the infraorbital nerve. With the exception of the infraorbital nerve, which is a branch of the maxillary division of the trigeminal nerve, all these nerves are derived from the first (ophthalmic) division of the trigeminal nerve. Innervation to the bulbar conjunctiva comes from the long ciliary nerve, which branches off the nasociliary nerve from the ophthalmic division of the trigeminal nerve.14 Most of the innervation to the superior palpebral and forniceal conjunctiva comes from the medial and lateral branches of the supraorbital nerves.1 The supratrochlear, infratrochelar, and lacrimal nerves innervate this region as well. The inferior palpebral and inferior forniceal conjunctiva receive sensory innervation from the lacrimal nerve laterally and from the infraorbital nerve medially.14 With the exception of some pressure sensation in the caruncle and the palpebral region, pain is the only sensation perceived on the conjunctiva.26
All nerves form a network in the conjunctiva and terminate either as specialized endings (e.g., end bulbs of Krause) or as free endings on blood vessels and epithelial cells. The majority of nerve endings in the conjunctiva are free and unmyelinated, forming a plexus in the superficial part of the substantia propria (Figs. 29.10 and 29.11). Many of these fibers end on blood vessels, whereas others form an intraepithelial plexus around the bases of the epithelial cells, sending free nerve endings between them.1
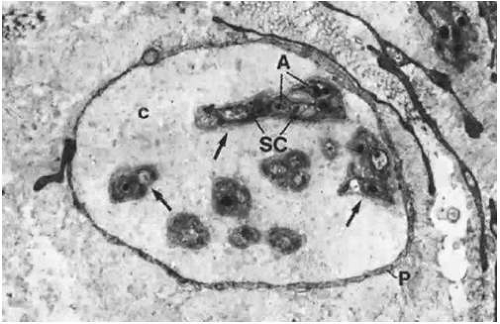 Figure 29.10. A nerve fiber bundle in the conjunctival stroma composed of several unmyelinated nerve fibers (arrows) surrounded by a layer of perineurium (P); intervening collagen fibrils also are present (c). Each unmyelinated nerve fiber is composed of axons (A) enwrapped with Schwann cells (SC) (×13,700). |
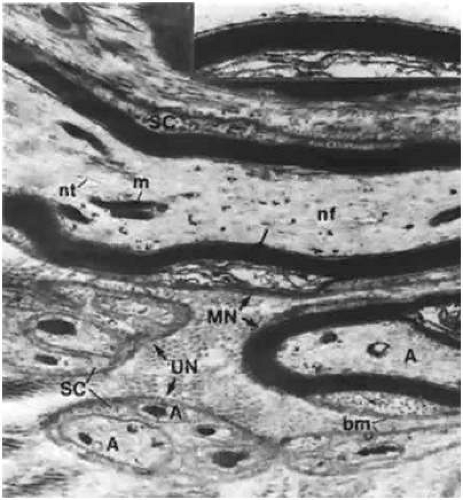 Figure 29.11. Substantia propria of the bulbar conjunctiva showing myelinated (MN) and unmyelinated (UN) nerve fibers. In both fibers, axons (A) are wrapped with Schwann cells (SC) that have a basement membrane (bm). However, the axons are single and have a myelin sheath (ms) in the former; they are multiple and have no myelin sheath in the latter. Also shown are the neurofilament (nf), neurotubule (nt), and mitochondria (m) (×20,800). Inset. Higher-power view of a portion (arrow) of the myelin sheath, showing the lamellar structure (×40,000). |
Also present in the conjunctiva are the end bulbs of Krause, which are specialized, compact nerve endings, 0.02 to 0.1 mm in length and surrounded by a connective tissue capsule. The bulb itself may be single with complex structure, or it may be compound. These end bulbs are relatively rare and vary in distribution. Their exact function remains unknown. It has been hypothesized that the end bulbs of Krause are really a stage in the growth cycle of specialized nerve end organs.27
Specialized Conjunctival Structures
The lacrimal caruncle and the plica semilunaris are two areas of specialized conjunctiva that are located in the nasal (medial) canthus of the eye. The caruncle is a small, flesh-like body that lies in the lacus lacrimalis on the medial side of the plica semilunaris. It is a part of the margin of the lower lid that becomes cut off by the development of the inferior canaliculus. The caruncle is covered by stratified, squamous epithelium similar to the skin, but it is not keratinized. Like the skin, it contains hair follicles as well as sebaceous and sweat glands within its subepithelial stroma.28 In contrast to the skin, the caruncle contains accessory lacrimal glands, such as Krause glands. The deep connective tissue in the caruncle is made up of parts of the septum orbitale and the medial check ligament. Numerous goblet cells can be found singly or in groups within the caruncle (Fig. 29.12). The blood supply of the caruncle comes only from the superior medial palpebral arteries. However, trauma or damage to the caruncle can cause profuse bleeding because of the abundant blood supply to this tissue.1 The caruncle is innervated by the infratrochlear nerve. Lymphatic drainage of the caruncle, along with the rest of the medial portion of the conjunctiva, flows into the submaxillary lymph nodes. In rare instances, cartilage may be present in the stroma.29
 Figure 29.12. Epithelium of the caruncle. It has many goblet cells (GC), some protruding toward the conjunctival sac. Small microvilli (mv) of the goblet cells also are seen (×12,200). |
The plica semilunaris is a semilunar fold of conjunctiva that lies lateral and slightly posterior to the caruncle in the medial canthus. It is more or less vertical, and its concave side faces laterally. Because the lateral border is free, a cul-de-sac of approximately 2 mm in depth is formed when the globe is adducted. It is practically nonexistent when the globe is abducted. The plica may correspond to the nictitating membrane in lower vertebrates. In humans however, it is likely a vestigial structure consisting of a lax fold of conjunctiva that allows full lateral movement of the eye. The plica consists of eight to ten layers of epithelial cells containing many goblet cells. Langerhans cells also may be present in the epithelium. The substantia propria is composed of adipose tissue and nonstriated muscle supplied by sympathetic nerves. The connective-tissue stroma of the plica is loose and highly vascular.
Conjunctival Glands
The conjunctiva and the adnexa contain many glands that are responsible for secreting the various components of the precorneal tear film. These glands are most abundant in the fornices and tend to become less frequent as one moves toward the limbus.30 In conjunction with the main lacrimal gland, the accessory lacrimal glands of the conjunctiva are responsible for producing the aqueous layer of the tear film. The accessory lacrimal glands are located in the connective tissue of the conjunctiva and include the glands of Krause and the glands of Wolfring.
The glands of Krause are located in the deep subconjunctival connective tissue of the upper fornix. About 42 Krause glands are located in the upper fornix, and approximately six to eight are located in the lower fornix. Krause glands also can be found in the connective tissue of the caruncle.26 The ductules of these glands join to form a single duct that opens into the fornix. The fine structure of the Krause glands is essentially the same as that of the lacrimal gland in the orbit. Clinically, it is important to preserve the integrity of the superior border of the upper tarsus during surgery, because inadvertent excision of these accessory lacrimal glands can cause a postoperative dry-eye problem.
The glands of Wolfring are accessory lacrimal glands that are larger than the Krause glands. Two to five glands of Wolfring are located on the upper lid, along the superior border of the tarsus.1 Two glands are present along the inferior edge of the lower tarsus. The excretory duct of each gland is lined with basal cuboidal cells, similar to the conjunctival epithelium onto which it opens.
A third type of conjunctival gland, the Manz glands, are located at the superior limbus. These glands are actually intraepithelial collections of goblet cells that produce mucinous secretions. Manz glands are located on the nasal side of the bulbar conjunctiva and have been identified in certain animals, such as the pig, calf, and ox, but they are thought to be absent in humans.1,31
Histology
Conjunctival Epithelium
Like other mucous membranes, the conjunctiva is composed of two layers: the epithelial layer and the substantia propria layer. The conjunctival epithelium is the stratified, nonkeratinized superficial layer of the conjunctiva. It is unique among other nonkeratinized epithelia in the body, because it contains goblet cells (Fig. 29.13). The fine structure of the epithelium varies from one region to another. At the mucocutaneous junction, the keratinized, stratified squamous epithelium of the lid margins quickly transitions into the moist, nonkeratinized squamous epithelium of the marginal conjunctiva. From there, the remaining palpebral conjunctival epithelium is composed of stratified cuboidal cells. Columnar cells are seen in the fornices, and stratified squamous cells make up the epithelium of the bulbar region (Fig. 29.14).32,33 It is believed that the predominance of the flatter squamous cell type around the limbus and the lid margin is due to an adaptation to increased mechanical pressure on the limbal and marginal epithelial zones.33
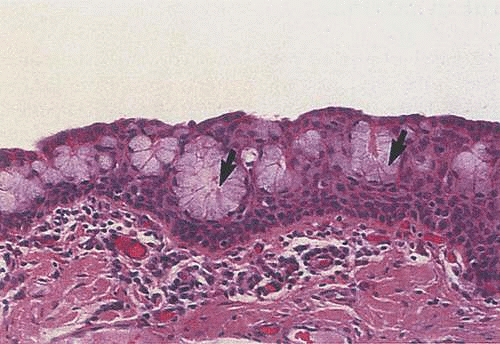 Figure 29.13. Goblet cells (arrows) are shown within the conjunctival epithelium. Scattered mononuclear cells also are seen within the underlying substantia propria (H&E ×66). (Reproduced with permission from Sternberg SS, ed. Histology for pathologists, 2nd ed. Philadelphia: Lippincott-Raven, 1997:321.) |
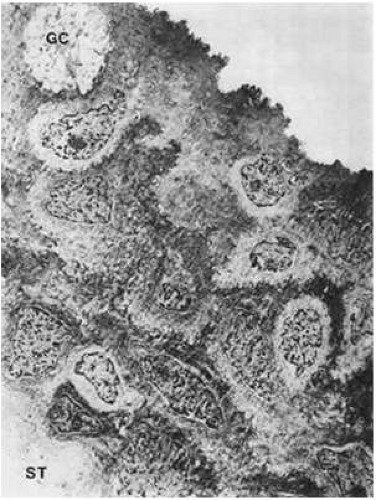 Figure 29.14. Bulbar conjunctival epithelium composed of irregularly piled polygonal epithelial cells. The surface is uneven and beset with microvilli. The basal line between the epithelium and the stroma (ST) is undulated. A goblet cell (GC) also is shown (×4,300). |
The thickness of the conjunctival epithelium has been reported to range from two to four layers in the tarsal conjunctiva, six to eight layers near the limbus, and up to 10 to 12 layers near the lid margins.34 It has been postulated, however, that the reported variations in the conjunctival epithelium thickness may actually be an artifact due to different degrees of stretching at the time of fixation and histologic examination.2
Overall, the conjunctiva has a relatively flat surface, except at the limbus and the tarsus, where the palisades of Vogt and the crypts of Henle are located, respectively.29 The palisades of Vogt (limbal palisades) are a radial arrangement of rete pegs and papillae formed from the conjunctival epithelium and stroma around the corneal periphery. The papillae radiate outward from the corneal edge into the conjunctiva, much like the spokes of a wheel.19 These palisades are present all around the cornea, but are most prominent superiorly and inferiorly. Small nerves, vessels, and lymphatics run the length of the papillae. The nerves are unmyelinated and branch considerably upon entering the conjunctival stroma and basal epithelial layer. The crypts of Henle are 0.5-mm appendix-shaped invaginations that are most prominent in the nasal half of the tarsal conjunctiva.35 They are tubular in shape and contain goblet cells.1
Considerable variation occurs in the appearance of the cells of the conjunctival epithelium. In a study examining the ultrastructure of the conjunctiva, Steuhl divided the conjunctival epithelial cells into five different types, based on their morphologic characteristics.33 According to this classification system, type I cells are goblet cells. Type II cells have numerous, electron-dense granules in the apical cytoplasm and are the most commonly seen cells of the epithelium. Type III cells possess well-developed Golgi complexes. Type IV cells are characterized by large quantities of rough endoplasmic reticulum, and type V cells are defined by their high content of mitochondria.
Functional Histology of the Conjunctival Epithelium
The histologic features of the conjunctival epithelium can be related to its many physiologic functions, which include protection and maintenance of the ocular surface. The barrier properties of the conjunctival epithelium are especially important with regard to ocular drug delivery. Intercellular spaces at the epithelial surface of the conjunctiva are closed by tight junctions at the apical ends of the superficial cells.36 This creates a semipermeable membrane at the ocular surface that facilitates the intercellular transport of lipid-soluble molecules but limits the passage of hydrophilic solutes.1,36 As a result, proposed mechanisms for enhancing the passive transport of ocular drugs across the conjunctiva include the development of more lipophilic agents and the use of enhancers to open tight junctions, thereby increasing paracellular penetration.37,38 The intercellular spaces of the conjunctival epithelium are wider than are those seen in the corneal epithelium.1 It has been reported that the pore radius of corneal epithelium is only 2 nm, whereas the pore radius of the bulbar and palpebral conjunctiva are 3.0 and 4.9 nm, respectively.39 Interepithelial openings of 1 to 3 μm, which represent the openings of interepithelial goblet cells, can be appreciated on scanning electron microscopy. Larger openings (10 to 60 μm) are due to the openings created by the epithelial rugae. Beneath the epithelial surface, extensions of the cytoplasm (microvilli) protrude into the intercellular spaces, where desmosomes occasionally connect neighboring cells.40 Microvilli also can be seen on the epithelial cell surface.40 Longer microvilli are found on the columnar cells of the fornices, while progressively shorter microvilli are seen on the squamous cells of the bulbar conjunctiva (Fig. 29.15A and B).41
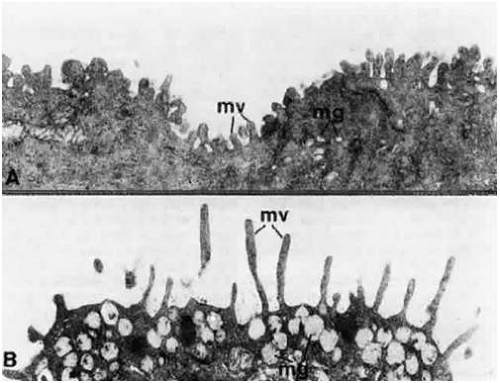 Figure 29.15. Transmission electron micrographs of the microvilli (mv) on the bulbar conjunctival epithelial surface (A) and the forniceal conjunctival epithelial surface (B), showing the difference in length. Microvilli are short in the former and long and slender in the latter. Mucous granules (mg) are seen in both images, with those in B exhibiting fibrillogranular contents (A. ×26,000; B. ×26,000). |
Perhaps the most important function of the conjunctival epithelium is its contribution to the tear film. Tears are a complex fluid that coat the ocular surface and serve multiple purposes. They act as a protective interface from the external environment, lubricate and maintain the ocular surface, and help preserve corneal transparency.5 The tear film is classically thought to be composed of three layers: (a) a superficial lipid layer; (b) an aqueous layer; and (c) an inner mucin layer. Each layer is secreted by a different gland or cell type, and the presence of each layer is crucial to the overall stability of the tear film. However, given the hydrophilicity and solubility of mucins, recent models suggest that the aqueous and mucin layers actually are intermixed, as opposed to being two sharply demarcated and separate layers in the tear film.42 Regardless, proper functioning of the conjunctiva is necessary for the formation of a healthy tear film, considering that the conjunctival glands help form the aqueous components while the goblet cells and epithelium secrete the mucins. Table 29.2 summarizes the components of the tear film.
TABLE 29.2. Properties of the tear film | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||
The conjunctival epithelium synthesizes a variety of high-molecular-weight glycoproteins called mucins. They can be categorized as either secretory or membrane-associated. Currently, over 15 types of mucins have been identified, with each being numbered in the order that it was discovered.42,43 Of these, conjunctival epithelial cells produce only three membrane-associated mucins (MUC1, MUC4, and MUC16).44,45 MUC2 and MUC5AC are secretory, gel-forming mucins that also are found in the conjunctiva.43 MUC2 is present in small amounts, and its relative distribution in the conjunctival epithelium is still uncertain.43,46,47 MUC5AC, on the other hand, is a secretory product of the conjunctival goblet cells and is a major component of the mucous layer of the tear film.48 It was believed that MUC1 was responsible for spreading the MUC5AC of the goblet cells along the ocular surface and anchoring the tear film.43 However, MUC1 knockout experiments in mice showed no change in the stability of the tear film.49,50 As a result, the specific role of MUC1 is an issue of some debate. Kardon and associates did show, however, that MUC1 may serve a protective role by preventing pathogens from adhering to the ocular surface.49 MUC4 is thought to contribute to the mucous layer of the tear film and, in fact, it is the only membrane-spanning mucin that has been found in tear samples to date.51 MUC16 was discovered recently, and little is known about its function.52 Further studies are needed to elucidate the exact role of all these mucins on the conjunctival surface.
The conjunctival epithelium is also unique in that it has the capability of both absorption and secretion. The epithelial cells are believed to be the primary source for fluid transport across the conjunctiva.5 In a study of patients with occluded efferent tear ducts, 30% of the technetium placed in the conjunctival sac was absorbed within 15 minutes.53 The conjunctiva’s ability to absorb water and electrolytes aids in the maintenance of the ocular surface by allowing it to adjust the concentration of various tear-film components.5 In addition to mucins, histochemical and immunochemical studies have shown that the conjunctival epithelium is capable of secreting multiple proteins and cytokines.54,55 Like the lacrimal gland, the conjunctiva is thought to produce antibacterial proteins, similar to lysozymes, that protect the ocular surface. Some evidence also suggests that the conjunctiva may secrete growth factors into the tear film.5
Cells of the Conjunctival Epithelium
The conjunctival epithelium is composed of multiple types of cells. Each cell type possesses a unique structure that allows it to carry out a specific physiologic function in the conjunctiva and ocular surface.
Apical Cells
The apical cells of the conjunctiva are located along the epithelial surface. These cells have a hexagonal shape and are arranged in clusters around a central core of cells, much like the petals of a flower (Fig. 29.16A and B). They range in size from 6 to 10 μm in the tarsal conjunctiva to 3 to 20 μm along the limbus.56 The apical cell surface is completely studded with microvilli and microplicae. The diameter and height of the microvilli are approximately 0.5 μm and 0.5 to 1 μm, respectively.1,57 They are separated by gaps of 0.5 to 1 μm.1,57 Microplicae, which are thought to be fused microvilli, are approximately 0.5 μm wide and up to 3 μm long.33 These structures are important not only for enlarging the surface area of the epithelium available for absorption, but also for stabilizing and anchoring the tear film.34,58,59 The tips of the microvilli and microplicae express mucin-like glycoproteins, which aid in the maintenance and spread of the tear film over the rest of the ocular surface.43,60 The conjunctival microvilli also have an important role in the ocular immune response because of their ability to absorb foreign particles during infection. The phagocytic ability of the conjunctiva has been confirmed by studies showing that conjunctival epithelial cells are capable of internalizing bacteria and even experimental latex microspheres.61,62 The high alkaline phosphatase activity of the microvilli also may aid in its antimicrobial function. Actin filaments within the microvilli help anchor the cell membrane to the cytoskeleton.63
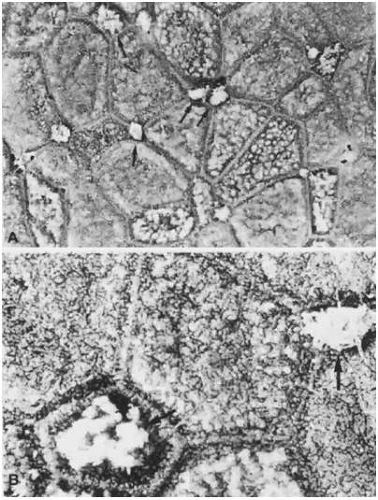 Figure 29.16. A. Low-power SEM of the epithelial surface of bulbar conjunctiva, showing relatively distinct cell borders. Polygonal cell shape and their petaloid arrangement can be appreciated. Large white spots (arrows) represent the mucous substance secreted through the openings of the goblet cells. The tissue was treated with 20% acetylcysteine for 10 minutes, but mucoid substance still remained in places on the epithelial surface. B. Higher-power SEM of the same epithelial surface shown in A. Microvilli cover the entire surface, but the cell borders are clearly distinguished from the adjacent areas because of different distribution densities of microvilli. Arrows indicate the surface of goblet cells (A. ×2,300; B. ×6,500). |
The appearance of these cells on scanning electron microscopy depends on the density, size, and distribution of the microvilli and microplicae. As a result, the apical cells of the conjunctival epithelium can be divided into three types by the amount of scattering they exhibit on electron microscopy: light-, medium-, and dark-colored.3,64 The appearance of the light-colored cells is attributed to a greater density of microvilli.43 Mucous production is greatest in light-colored cells and, as a result, mucin expression appears to be a function of microvilli and microplicae density.3,43 Medium- and dark-colored cells are thought to have a lower density of microvilli than do light-colored cells.43 In addition, their microvilli are shorter and thicker in comparison. The most superficial cells with the least microvilli are believed to be the oldest cells of the ocular surface.43
Intracellularly, the apical cells contain numerous vesicles within their cytoplasm (Fig. 29.17). These structures also are known as subsurface vesicles. The vesicle contents stain readily with Alcian blue and periodic acid-Schiff, which suggests that they are glycoproteins. As a result, the subsurface vesicles are thought to deliver a transmembrane mucin product that may contribute in some way to the inner mucin layer of the tear film.2 Studies have shown that apical cells are capable of producing all three types of membrane-associated mucins found on the conjunctival epithelium: MUC1, MUC4, and MUC16.44,45,51,65,66 The importance of proper mucin production cannot be overstated. The hydrophilic, negatively charged nature of these mucins is critical in providing an effective barrier to potential pathogens, as well as in lubricating the epithelial surface.2 Table 29.3 summarizes the properties of the conjunctival surface mucins.
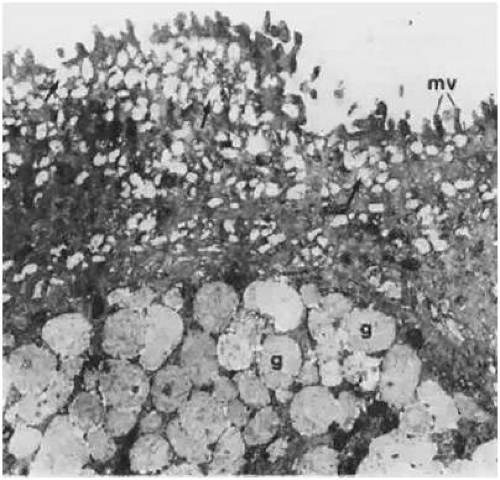 Figure 29.17. Superficial layer of bulbar conjunctival epithelium showing microvilli (mv) and numerous mucous granules (arrows) within the apical cells. Goblet cell vesicles (g) also are shown toward the bottom of this image (×24,700). |
TABLE 29.3. Mucins of the conjunctiva | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||
Basal Cells
The basal cells of the conjunctiva are relatively quiescent cells located deep to the apical cells and above the basal lamina. They attach the remaining epithelium to the underlying basement membrane via hemidesmosomes. Organelles, such as mitochondria and Golgi bodies, are relatively sparse and, when present, tend to be seen around the perinuclear area.1 The immediate periphery of the nucleus is free of organelles. This arrangement gives the appearance of a perinuclear halo around a large, electron-dense nucleus.1 It has been postulated that basal cells of the conjunctival epithelium differentiate into mucin-producing goblet cells, because immature goblet cells have been observed among other conjunctival epithelial cells along the basement membrane.67
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


