(1)
University of Sydney, Sydney, Australia
Overview
Color is a subjective sensory phenomenon, not a physical attribute of an object.
Color perception arises from stimulation of cones by light.
Color perception varies with:
(a)
The spectral composition of light reflected from object
(b)
The ambient light surrounding the object
(c)
The subject’s level of visual adaptation
Humans can distinguish possibly 7–10 million colors [1].
Color and Light
Monochromatic light is colored light of a single wavelength (Table 24.1).
Table 24.1
Wavelengths corresponding to spectral colors
Spectral color
Wavelength (nm)
Violet
430
Blue
460
Green
520
Yellow
575
Orange
600
Red
650
A wide range of colors can be reproduced by an appropriate combination of the additive primary colors: blue, green, and red.
Complementary colors are two appropriately selected colors which mix to produce white light.
Metamers are physically distinct combinations of light that appear identical; e.g., monochromatic yellow light is a metamer of yellow produced by red and green light combined.
Perception of Colors
Colors can be subjectively appraised and graded by three qualities: hue, saturation, and brightness.
(i)
Hue
Hue is the aspect of color allowing it to be assigned a position on a color spectrum.
It is related to the wavelength of monochromatic light.
In paint theory, hue is often referred to as a “pure color.”
(ii)
Saturation
Color saturation is determined by dilution of hue by white.
Pure hue is complete saturation; it can be progressively desaturated until white is reached.
(iii)
Brightness
Brightness is the apparent intensity of color: varying from very dim to dazzling.
It is ‑related to the object’s radiant energy.
Phenomena in Color Perception
1.
Colour inconstancy
An object’s apparent color changes by altering background spectral composition (Fig. 24.1).

Fig. 24.1
Color inconstancy. The inner square is identical on either side of the image. It appears pale blue against a deep orange background; it appears darker against a pale blue background
Similarly, the color can appear to remain the same despite changes in ambient light effecting the spectral composition of light from the object and its background [3, 4].
This is because color perception is not due to the absolute spectral composition of light from an object, but the spectral composition relative to the background.
2.
Trichromacy: Cone Transmission of Color
Normal color vision is trichomatic, mediated by three types of cone receptor distinguishable by their spectral sensitivity:
(a)
Short-wavelength-sensitive (SWS or S) cones
(b)
Middle-wavelength-sensitive (MWS or M) cones
Each type has a distinctive photoreceptor pigment that determines spectral sensitivity.
There is considerable overlap in spectral sensitivity between the three cone populations; however, each has a specific peak spectral sensitivity (Table 24.2, Fig. 24.2).
Table 24.2
Spectral sensitivity of three types of cone receptors
Cone population
Spectral sensitivity peak (nm)
Major color sensitivity
Short wavelength sensitive (S)
440–450
Blue
Middle wavelength sensitive (M)
535–550
Green
Long wavelength sensitive (L)
570–590
Red
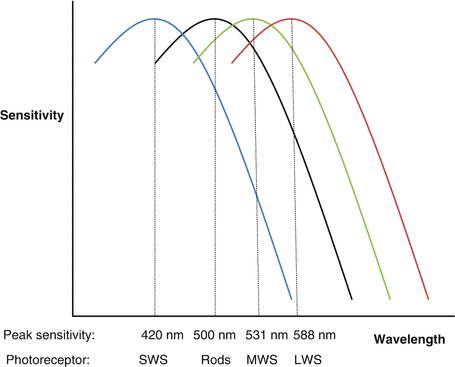
Fig. 24.2
Overlapping spectral sensitivity curves for SWS, MWS, LWS cones and rods
The wavelength of light determines the likelihood of stimulating each cone type.
Most cones are either M or L; S cones make up 5–10 % and are not found within the central fovea [10].
Trichromacy allows a full range of colors to be distinguished [11].
Opponent Processes: Color Processing in the Inner Retina and Lateral Geniculate Nucleus
The three cone types give rise to perception of hues arranged in two opponent pairs:
(a)
Red/green (R/G)
(b)
Blue/yellow (B/Y)
Opponent processing is found in inner retinal circuitry and the lateral geniculate nucleus.
1.
Inner retinal color processing
Inner retinal color processing occurs through distinct R/G and B/Y opponent channels.
(i)
Red/green opponency
R/G perception is conveyed by color opponent midget ganglion cells (MGCs) with center–surround antagonistic receptive fields (CSARFs) [12, 13].
Color opponent midget cell CSARFs are organized such that the center and surrounds are dominated by opposing M and L cone types; i.e., M–center/L–surrounds or L–center/M–surrounds.
(ii)
Blue/yellow opponency
B/Y opponency is conveyed through small bistratified ganglion cells that receive:
(a)
ON signal from S cone inputs (the blue signal)
Other combinations of S cone input with M and/or L cone inputs are reported [19] but not yet well understood.
2.
Lateral geniculate nucleus (LGN) color processing
Color opponent LGN cells are parvocellular cells in laminae 3–6 that receive MGC projections [22].
They have similar receptive field properties to the MGCs that provide their input.
Most R/G parvocellular cells transmit color opponency; these have CSARFs which have color opponency to large spot sizes and spatial luminance sensitivity (acuity) to small spots [23].
Some koniocellular LGN cells receive small bistratified ganglion cell B/Y opponent information [24].
Color Processing in the Visual Cortex
1.
The primary visual cortex (V1) (see Chap. 14, The Primary Visual Cortex)
Chromatic projections arrive in V1 along separate LGN R/G and B/Y channels [16].
Information from parvocellular channels projects to V1 layers 2 and 3; parvocellular projections are used for both achromatic luminance sensitivity and R/G color processing [25].
B/Y signal is conveyed via koniocellular channels that project to superficial layers of V1 [16, 24].
2.
Color-sensitive neurons in V1
Sensitivity to color in V1 occurs predominantly through the combined activity of two kinds of neurons: single–opponent and double–opponent cells.
These have distinct functions: the single-opponent cells respond to large areas of color, while double-opponent cells respond to color boundaries, patterns, and textures.
(i)
Double-opponent cells
(ii)
(iii)
Complex opponent cells
3.
The extrastriate visual cortex (see Chap. 15, The Extrastriate Cortex)
Get Clinical Tree app for offline access

(i)
V2
Neurons in the cytochrome oxidase (CO) blobs of V1 send projections to color-selective neurons in the thin stripes of V2 [36, 37].
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree

