Fig. 28.1
(a) During normal development, sensory neuroblasts delaminate from the otocyst epithelium and migrate through the periotic mesenchyme (POM). During this phase there is a period of proliferation called transit amplification. The cells then aggregate and start to differentiate. The process occurs over a period of approximately 4 days and migration occurs over a distance of up to 500 μm. (b) Transplanted precursor cells of auditory neurons (N33) recapitulate the process of migration, aggregation, and morphological differentiation in the adult rat auditory nerve tract. RC Rosenthal’s canal, SGC spiral ganglion cells, HC hair cells (cited from Sekiya et al. [6])
28.3 Neurite Extension, a Prerequisite for Auditory Nerve Regeneration
One crucial prerequisite in the study of auditory nerve regeneration is that donor cells extend definite neurites like default bipolar shape or at least two from multipolar processes of donor cells, toward hair cell and cochlear nucleus regions. Without such elongated neurites, synaptic formations could not be expected to be formed at both sides of transplanted cells.
To promote neurite extensions from donor cells, growth factors are indispensable. Donor cells need two different nutrient sources for survival and differentiation; one is peri-grafting and the other is post-grafting sources. In peri-grafting period, usually there is no other alternative but to provide immediate nutrients externally, such as donor cells transplanted with nutrient-rich medium. In post-grafting period, donor cells should settle down in a location where nutrients such as neurotrophic factors are constantly and permanently provided (see Chap. 7).
It cannot be expected that sustained supply of neurotrophins is provided to donor cells in the scala tympani [2, 13, 14], because the composition of perilymph is similar with the cerebrospinal fluid that hardly contains an ample amount of growth factors [15]. To compensate such a drawback of perilymphatic fluid, co-grafting of growth factor-producing cells or infusion of growth factors to transplanted site was performed with success [2, 13]. Neuronal differentiation and survival rate of mouse ES cells transplanted into the scala tympani were greatly enhanced only when GDNF was continuously infused into the scala tympani [2]. When ES cells were transplanted into the scala tympani with NGF-producing dorsal root ganglion (DRG) cells, the transplanted cells extended neurite-like processes toward the host SGCs and their peripheral nerve processes [13].
28.4 Auditory Nerve as an Avenue for Donor Cell Migration/Differentiation and Route for Trophic Factors Supply
Emerging evidence indicates auditory nerve trunk is more favorable as a site for cell transplantation than any other places in attempting auditory nerve regeneration [6, 10, 16–19]. It is likely neurotrophins synthesized in hair cells directly flow down into auditory nerve and “showering” of these factors on transplanted cells within auditory nerve trunk enhances their migration and neurite elongation. ES cells after neural induction had been applied were transplanted into the basal portion of auditory nerve. Four weeks after transplantation, the donor cells extended neuritic processes markedly both peripherally and centrally [16]. ES cells converted to neural progenitor cells were infused into auditory nerve trunk in deafened gerbils. By 2–3 months, the donor cells had extended neurites abundantly into the organ of Corti through empty Rosenthal’s canal (RC) [17]. A study demonstrated auditory precursor cells transplanted into auditory nerve trunk extensively migrated both distally and proximally [6]. ES cells placed at the internal auditory meatus portion of the auditory nerve that had been previously injured by compression migrated along the auditory nerve, passed through empty RC and the habenula perforata, and finally reached the scala media, a suitable place to form the synapse with hair cells, indicating the transplanted cells can reach there without breaching the membranes sealing the peri- and/or endolymphatic spaces [10]. These results indicate auditory nerve can be used as a useful avenue for donor cell migration/differentiation and a route for trophic factor supply, especially in auditory neuropathic-type auditory nerve degeneration where hair cells are intact (see Chap. 7).
There were reported various cell transplantation techniques into the cochlea [5]. To directly reach the CNS portion of auditory nerve, it is necessary to enter into intracranial space (Fig. 28.2) after incising single membrane, the dura mater [6, 20]. In trans-round window/perilymphatic space approach, one more membrane sealing the scala tympani has to be incised before the dura mater is incised that lines the inner surface of internal auditory meatus within which auditory nerve trunk resides (Fig. 28.2). Breaching of the membrane sealing perilymphatic spaces induces inevitable disturbance of the homeostasis of the inner ear fluid environment, thus placing hearing at risk [21–23].
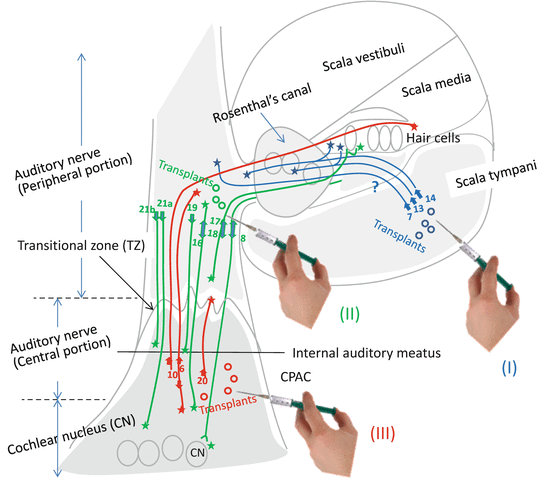
Fig. 28.2
Reported results of cell transplantation experiments to rebuild lost auditory neurons. The reports in which neurite extensions were confirmed are mainly listed. (I), (II), and (III) indicate routes of cell transplantation. (I) scala tympani approach (blue); (II) trans-round window/perilymphatic space approach (green); (III) retromastoid auditory nerve approach (red). Numbers denote reference numbers. Trajectories of cell migrations (blue, green, and red curved lines) are not necessarily confirmed in all cases as is representatively shown by a question mark. One sided and bidirectional arrows indicate initial transplanted sites and stars farthest point that donor cells reached. See the detailed results in the text. CPAC cerebellopontine angle cistern
28.5 Commitment of Auditory Neurons for Regeneration in Cochlear Nucleus Cells
In a study with embryonic DRG neurons cocultured with brainstem slice, numerous neurites from DRG neurons extended toward cochlear nucleus (CN) region, indicating an endogenous attractive factors were released from the cochlear nucleus [24]. Survival of cochlear nucleus cells intimately depends on the auditory neurons; innervation by auditory neuron and subsequent synaptic activity are essential for normal development of the cochlear nucleus cells [25]. With reinstalling of auditory nerve spike activities using a cochlear implant in deafened cats, the endbulb of Held was rescued [26]. These results indicate regeneration of auditory neurons is prerequisite for regeneration in CN and related structures.
A study showed statoacoustic ganglion cells transplanted in the auditory nerve migrated into the CN in the animals deafened with beta-bungarotoxin [20]. DRG neurons and ES cells transplanted in transected auditory nerve migrated close to ventral cochlear nucleus [21]. Similarly, auditory precursor cells transplanted into the auditory nerve migrated not only peripherally but also centrally [6]. However, one potential dilemma in this regard is that the more proximal the donor cells migrate toward the CN region, the more they become remote from the hair cell region and vice versa. It has not been clarified yet whether single donor cell can bridge between hair cell and cochlear nucleus cell as in default condition or multiple neurons with interneurons are needed as was demonstrated in a study where severed pyramidal tract fibers were reconnected using multiple neurons [27].
28.6 Is “Massive” Regeneration of Spiral Ganglion Cells Needed for Hearing Restoration?
Studies indicate that there is no significant correlation between clinical performance of cochlear implant (CI) and the residual absolute number of auditory neurons and hence it is believed that some other unidentified clinical variables are related to CI performance [28–31]. Theoretically, however, some minimal number of auditory neurons should be left functional. In one postmortem study of patients with CI, the least number of the SGCs of the patient was 1,443 that is approximately less than 5 % of total number of SGC, 30,000 [31]. Coincidentally, it is reported that preservations of axons and myelination are not proportional to the degree of neurological recovery in spinal cord injury [32] and effective locomotion was found to recover when only of 5–10 % of the original axonal population was maintained [33]. Based on these observations, there seems to be a possibility even if regenerated auditory neurons were small in number, its effect would be significant to patients’ hearing. To clarify this important point, morphological studies corroborated by functional evaluation are needed in our future experiments [8].
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


