Binocular Vision
Christopher W. Tyler
Eyes come in pairs, providing special capabilities that are not available to a single imaging system. This chapter provides an overview of the ways that visual processing has evolved to utilize the joint capabilities of a dual optical system for the perception of the third spatial dimension, of the visual deficits that can occur specific to the binocular coupling of this system, and of current approaches to ameliorating those deficits.
PHYLOGENETIC BACKGROUND
It is remarkable that virtually all animals have their eyes arranged in pairs, despite the tremendous variety in general morphology across species.1 Cyclops are essentially unknown outside mythology. Over many types of image-forming structures, from the pigmented eyespots of the flatworm, the pinhole eyes of the nautilus, and the compound eyes of insects, to the familiar mammalian arrangement, all seem to favor a paired organization. Even among invertebrates, examples of multiple eyes (arachnids, crustaceans) usually consist of two major eyes, with the subsidiary eyes lacking image-forming capability. The reasons for this dominance of binocular vision are difficult to establish, but five possibilities may be suggested.
The second eye could be a safety factor against injury or disease, improving the chance of retaining visual capability when part of the visual field (a scotoma) or the whole eye is damaged.
Two eyes may be used to achieve a 360-degree field of view, because optics are limited to approximately 180 degrees (in contrast to the compound eyes of insects, which have no inherent limit, but nevertheless always occur in pairs).
Where the fields of the two eyes overlap, there is an advantage in detection of faint images and rejection of the optical distortions within the eyes.
The presence of two mobile eyes allows the organism to converge the line of sight on distant objects and obtain a reading as to their absolute distance.
And, probably the most dramatic reason for binocular vision, two eyes permit stereoscopic depth perception–the ability to use the differences in the images caused by each eye viewing from a slightly different viewpoint, known as binocular disparities, to perceive distance in the third dimension of visual space.
Presumably all animals develop a visual system with some weighting among these different factors. Most lower animals take advantage of the possibility of a 360-degree field of view by having the eyes pointing in opposite directions. This advantage applies to fish, many birds, and to a large extent, mammals that are preyed upon. However, most predatory animals, from spiders and crustaceans through birds to mammals and humans, tend to have the eyes facing in the same direction. This arrangement is, presumably, to take advantage of stereoscopic depth perception, which has been demonstrated in humans, monkeys, cats, and falcons, and is likely to be widespread across other species.2,3,4,5
Actually, almost all animals show some degree of binocular overlap, even those with a full 360-degree field.1 The binocular overlap is almost exclusively in the forward direction (although it may be oriented upward or downward in different species). It is unclear, however, whether the binocular field is used to take advantage of stereoscopic depth cues or merely for the improvement in image reliability by binocular comparison. Certainly most birds have a region of improved visual acuity in the binocular segment in addition to the laterally directed monocular foveae (Fig. 1). They, therefore, have sufficiently good visual acuity to use binocular disparity cues for stereoscopic depth perception.
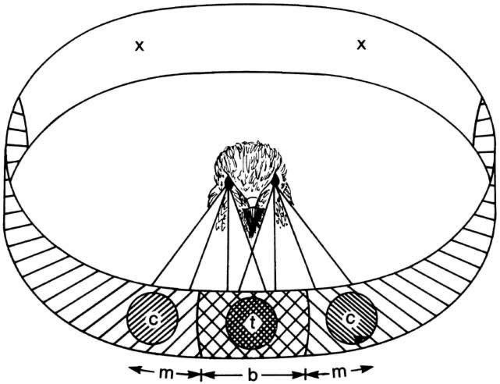 Fig. 1 Projection of the three regions of high acuity in the visual field of a hawk. The central foveae in the two eyes project to two monocular regions (c), while the temporal foveae in each eye project to a binocular region (t). In the case of this species, a rear portion of the field is out of view (x). (Walls GL: The Vertebrate Eye. New York: Hafner, 1967) |
The relative unimportance of convergence angle as a primary cue to distance may be suspected by noting that many species have little or no convergence capability. This observation is true for many birds and, in particular, the owl, with its large binocular fields.6 A similar situation occurs in lemurs and would imply that convergence works mainly to optimize the alignment of the retinas for the assessment of binocular disparities for stereopsis rather than as a primary cue to distance. Still, some animals may use the muscular convergence information that allows minimization of disparity by aligning the eyes to help determine the distance of an object.
In addition to the benefits of binocular vision, any animal with two eyes obtains the advantage of duplication against dysfunction; if duplication were a principal advantage, however, a further proliferation of the number of eyes might be expected.1 The advantage obtained against dysfunction is, therefore, probably of minor importance. We are left with the conclusion that the major reasons for binocular vision are probably to achieve a large visual field and to enable the use of stereoscopic depth perception.
SENSORY ASPECTS OF BINOCULAR VISION
With the two eyes aligned on a stimulus field, there are several sensory aspects of binocular vision that arise from the relative configurations of the stimulus details. These will be discussed on the assumption that the eyes stay aligned on a defined fixation target. Specifically, the alignment is defined by the angular direction of the region of highest acuity, the fovea. The orientation of the globe is referenced to this control region of the retina, centered at the back of the eye; angular distance from the fovea in any direction is known as eccentricity for targets in the visual field. In practice, of course, the eyes normally move to a variety of fixation positions both laterally and in depth as the observer moves the fovea to inspect a visual scene. However, the sensory processes need to be understood in terms of the effects of stable binocular presentation before the dynamic effects of moving eyes can be taken into account.
VISUAL DIRECTION
With the foveal fixation on one point, all other points in the visual field of one eye are seen as having different visual directions with respect to the fixation point. This two-dimensional spatial layout of visual directions is a defining property of the visual sense (as contrasted, for example, to the sense of smell, in which there is no spatial organization). If a stimulus point in space projects so as to have the same perceived visual direction in space when viewed with either eye, its image is considered to fall on retinal points that are exactly corresponding in the two eyes, as depicted in Figure 2. Departures from correspondence in the form of small differences between the image positions in the two eyes are termed binocular disparities (Fig. 3).
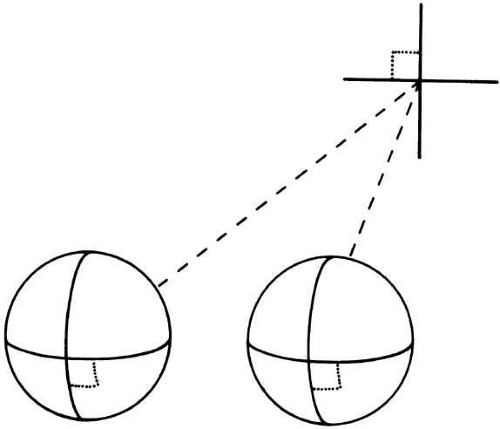 Fig. 2 Geometric retinal correspondence. When the eyes are aligned and viewing at infinity (shown here in perspective), a point at a given distance vertically and horizontally away from the fixation point (dashed lines) projects equivalent distances horizontally and vertically away from the foveae in the two eyes. These two points are then in geometric correspondence. |
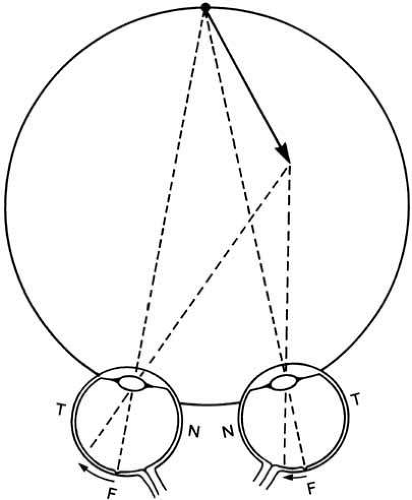 Fig. 3 Horizontal binocular disparities arise from objects (e.g., arrow) at different distances and give rise to stereoscopic depth perception. Here the arrowhead has a greater eccentricity on the temporal (T) retina of the left eye than on the nasal (N) retina of the right eye. Fixation is with the fovea (F). |
SENSORY FUSION
When there is a small binocular disparity in visual direction in one eye relative to the other, the binocular image still will be seen as a single fused image. The range of disparities for which binocular fusion occurs is known as Panum’s area. When the images are fused, the two images combine into a joint percept that resembles the one seen by each eye alone. Beyond this region of fusion, images are seen doubled or in diplopia, such that the separate images for the two eyes are both perceived in their true monocular locations.
DICHOPTIC STIMULATION
Images that are very different at corresponding retinal locations in the two eyes (e.g., images that derive from the same stimulus but are shifted, rotated, or magnified so that dissimilar contours are present in corresponding locations) provide dichoptic stimulation conditions. If such dichoptic images are of high contrast, then a binocular rivalry will be set up between the two monocular images, so that the observer is aware alternately of the image to one eye and the image to the other. If one eye is strongly dominant as a result of either stimulus characteristics or organismic variables, perception of the image in the other eye may be entirely absent because of interocular suppression.
If, however, the stimulus contrast is low for dichoptic stimuli, prolonged periods of dichoptic summation may be obtained, during which the different stimuli in the two eyes appear to be summed together as if their contrasts were added linearly throughout the dichoptic field. Dichoptic summation also is obtained for high contrast stimuli when the presentation time is brief (<150 msec).
STEREOPSIS
If two similar images are presented to the two eyes with a binocular disparity that has a horizontal component, the combined image will be seen in vivid depth nearer or farther than the point of fixation (Fig. 3) within some range of limiting conditions. Stereoscopic depth from horizontal disparities is perceived both in the region of binocular fusion of the monocular targets into a single image and also in the region of diplopia, where the image appears doubled but clearly at a different depth from zero-disparity targets. Stereoacuity is the smallest disparity interval that produces reliable depth discrimination under particular conditions.
DEVELOPMENT OF BINOCULAR VISION
NEUROPHYSIOLOGY OF DEVELOPMENT
The fundamental anatomic basis for binocular vision in mammals is the partial decussation of the optic nerves of the two eyes to bring the retinal information from corresponding areas of each eye together at the cortex, as considered in more detail in Figure 15. From the point of view of a neuron in the cortex, the region of the retina in which it responds to changes in local illumination is called the receptive field of that neuron. Most neurons in the cortex have identifiable receptive fields in both eyes, either at corresponding locations or close to them. At birth, the retina and optic pathway are not completely developed; however, the basic receptive field organization of neurons and cortical architecture are present.7 Neurophysiologic studies have demonstrated that the properties of neurons in the visual cortex are markedly influenced by visual experience during the first few postnatal months. Specifically, these developing neural properties have been determined to involve:
Ocular dominance, the relative strength of response from a particular neuron for input from the two eyes
Orientation specificity, the degree of tuning of the response to one specific orientation of a line stimulus
Binocularity, the degree to which the binocular response exceeds the response to stimulation of each eye alone
Disparity specificity, the degree to which the response is tuned to a specific binocular disparity of the stimuli in the two eyes
Note that the disparity specificity can be tuned to zero disparity, near disparity, or far disparity.
The excitatory connections of receptive fields located in both retinas or retinotopic projections are largely present at birth.8,9 However, simultaneous occurrence of patterned visual input to both eyes during the developmental period is necessary to maintain their association.10,11,12,13,14 For each binocular visual function, there is a period of sensitivity soon after birth during which the formation of receptive fields and other indices of visual processing may be disrupted unless full binocular visual input is available. The period of sensitivity to binocular disruption, produced by patching one eye of an animal so that there is only monocular visual input, is longer in monkeys than in cats. One index of binocular function is a susceptibility to ocular dominance shifts between the receptive fields of the two eyes. Such cortical dominance shifts seem to start at birth15,16 and show susceptibility up to approximately 1 year.11,12 Binocular vision could be affected by monocular deprivation up to 2 years of age.13,14
In the developing visual system, patterned visual stimuli seem to act not only as a catalyst but also as a guide in the consolidation, maintenance, and refinement of the neuronal connections of the visual cortex.15,16,17,18,19,20 Any disruption to the normal developmental conditions, such as a congenital or early-onset ocular misalignment or significantly subnormal vision in one eye, can prevent the eyes from developing the normal functional interrelationship.21
A possible function of this plasticity in neuronal properties during early visual experience might be to allow the opportunity to match the properties of feature-detecting cells to the most common features in the animal’s visual world, maximizing its capacity for analysis of the more important components of the environment.22,23 Also, the ability to make modifications in optimal disparity and the preferred orientation would ensure that binocular cortical cells adopt similar receptive field positions and preferred orientations of the two retinas. Both in immature visual systems and in binocularly deprived visual systems, the binocular responses of cortical neurons are observed to tolerate a wide range of retinal disparity, whereas visual experience narrows this range. Thus, early plasticity seems to be vital to the formation of cells with closely matched receptive field properties in the two eyes, which is a necessary condition to form the substrate for stereoscopic vision. Blakemore and Van Sluyters9 propose that “genetically specified, predominately monocular, simple neurons initially provide a ‘conditioning’ input to future complex cells and entrain them to respond to the same orientation. Both cell types ultimately gain matched input from the two eyes.”
There are well-established postnatal morphologic changes in the retina and retinal projections relative to the presence or absence of visual stimuli. In the lateral geniculate nuclei, neuronal cell growth is greatly reduced in the laminae with connections to a visually deprived eye as compared with a nondeprived eye.10,23 In studies in which one eye was deprived of light for 3 months after birth and was then allowed visual stimulation while the other eye was deprived, the difference in cell size between the lateral geniculate nuclei laminae is decreased and even reversed.24 The two types of retinal ganglion cells and geniculate cells seem to be differently affected by such deprivation.25,26 The large cells of the binocular segment (which are found mainly in the peripheral field projections) are much more affected than either the large cells of the monocular segment or the small cells found mainly in the projection of the area centralis.
Most of the cells of the superior colliculus of the midbrain are binocularly driven. Visual deprivation in kittens has consequences in the superior colliculus that can be considered to reflect changes in the visual cortex and, hence, in the corticotectal projection that is presumed to provide binocularity. The results of monocular deprivation experiments suggest that there is functional competition for effective synaptic input from the two eyes dependent on the actual firing pattern of afferent synaptic fibers, and that this competition represents a crucial process underlying the course and nature of the development of the visual system.11,12,13,19
The physiologic deficits observed in animals suggest a basis for understanding various human perceptual disorders. Monocularly deprived kittens, for example, do not develop the normal proportion of binocularly driven cortical neurons if the monocular deprivation is imposed during a specific stage in their maturation, which has been determined to be 4 through 12 weeks.27,28 Also, misalignment of the visual axes (which results in discordant binocular input) interferes with the development of binocular neurons when experimentally induced during this same period.19,29 Orientation specificity and disparity specificity of the binocular receptive fields for cat cortical neurons are similarly dependent on visual experience in that early postnatal period.17,18,19,29,30,31 Between the ages of 4 and 10 weeks, 1 week of monocular deprivation is sufficient to produce substantial degradations in stereopsis (as assessed by the difference between monocular and binocular performance on a jumping stand).33
NORMAL BINOCULAR DEVELOPMENT IN HUMANS
Similar interocular modifications effects have been observed in human development. Four main indices of binocularity have been used with human infants:
Accuracy of alignment of the visual axes of the two eyes;
Looking preferences by the infant for fixating stimuli containing a binocular disparity over flat stimuli;
Binocular facilitation of the visual evoked potential (VEP) amplitude for binocular stimulation to greater than the linear sum of the two eyes’ separate signals;
VEP responses to changes in binocular correlation and binocular disparity in dynamic random-dot stimuli that are designed to produce no changes that are visible monocularly.
Alignment of the eyes at birth is often imperfect, with exotropia (deviation away from the nose) occurring in up to 50% of newborns.33A Irregular fixations and alignment continue for up to 2 months. The eyes should be accurately aligned on targets of interest after this age, however.
Both preferential looking studies34,35 and VEP studies36 suggest that the earliest evidence of stereoscopic depth discrimination based on disparity alone occurs at approximately 3 months. What is remarkable about this capability is the rapidity with which it improves, with most infants being able to discriminate as little as 1 arc-minute of disparity by 5 months (see Fig. 4). The same studies show that near-disparity discrimination consistently developed earlier than the far variety. Several techniques37,38,39,40 have been used to track the development of fine stereoacuity in persons between the ages of 2 and 6 years. Although there are discrepancies up to a factor of 4 at any given age, all agree that stereoacuity improves gradually during this period to close to adult levels by age 6.
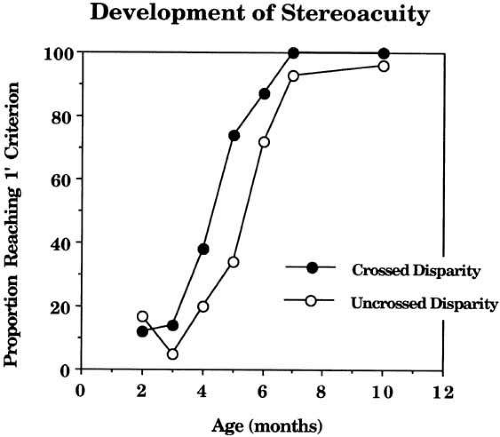 Fig. 4 Development of stereoacuity. Proportion of infants reaching 75% correct performance on a depth discrimination of 1 arc-minute of crossed (filled circles) or uncrossed (open circles) disparity. Note that the crossed disparity performance precedes the uncrossed by approximately 1 month. (Birch EE, Gwiazda J, Held R: Stereoacuity development for crossed and uncrossed disparities in human infants. Vision Res 22:507, 1982) |
The amplitude of the binocular VEP is about the same as the monocular VEP at birth, implying independent processing of the two eyes. An increase in the amplitude of the VEP for binocular relative to monocular viewing is termed binocular summation; doubling of the amplitude is termed full binocular summation and an increase beyond the range of doubling constitutes binocular facilitation. The degree of binocular summation of the VEP increases markedly into the facilitation range at approximately 3 months and then returns to adult levels of less than full binocular summation by approximately 12 months.41 This early period of enhanced binocular response appears to coincide with the period of binocular consolidation in humans, which is between 3 and 7 months of age, based on behavioral evidence (see Fig. 4). Correlation-specific VEP responses are not obtained before 2 months of age but are seen in most infants by 3 or 4 months.42,43 Thus, the two VEP indices are in fair agreement that the onset of binocular function in human infants occurs at approximately 3 months of age. There is a longer critical period for binocular misalignment in which there is some recovery of binocular function when the eyes of strabismic infants are realigned up to approximately 2 years of age.44,45,46
ABNORMAL DEVELOPMENT OF BINOCULARITY IN HUMANS
Binocularity and Eye Alignment
The perceptual appreciation of stereopsis involves the analysis of spatially disparate retinal signals from each eye under the condition of binocular single vision. However, about 4% of them are unable to use these disparity cues and perhaps another 10% who have difficulty in judging the type of disparity (i.e., near or far) that they perceive.47
One approach to the study of the presence and proportion of binocular cortical neurons in humans has involved the interocular transfer of orientation-specific aftereffects. That is, after adaptation with only one eye, the relative strength of the aftereffect when viewing with the unadapted eye is used as the measure of the degree of interocular transfer. If this interocular transfer is mediated by binocularly innervated cortical neurons, then the extent of the transfer should be an index of the proportion of cortical cells that are binocular.
Some reports involving both the tilt and motion aftereffects have noted a high correlation between the amount of interocular transfer from the adapted to the nonadapted eye and the observer’s level of stereoacuity. Among the individuals tested in both types of study were stereoblind subjects who exhibited little or no interocular transfer as well as individuals with a range of levels of stereoacuity from poor to normal.48 The magnitude of the correlations was 0.86 for the tilt aftereffect and 0.75 for the motion aftereffect. This association suggests that both stereopsis and interocular transfer depend on the presence of binocular neurons.
The absence of binocular cortical neurons could be the result of an early impediment to normal binocular vision resulting in their functional loss, comparable with that demonstrated in kittens, or alternatively it could be the result of a congenital deficiency. Congenital anomalies of the visual pathway, such that fibers from noncorresponding retinal areas come together in visual cortex, are known to occur in animals such as the Siamese cat and other albino mammals.49,50 Evidence for a similar misalignment in albinotic humans has been obtained from extensive VEP studies.51
Other studies have shown that a childhood history of strabismus does not eliminate interocular transfer.52,53 Two studies involving individuals with an early history of strabismus have used the interocular transfer phenomena to delineate the sensitive period in humans for the development of binocularity. Hohmann and Cretuzfeldt (Fig. 5) investigated the association between the levels of interocular transfer and the age at onset of the deviation and concluded that the end of the human critical period was approximately at 2 to 2.6 years of age.53 Additional support for this delineation of a human critical period for the development of binocularity54 suggest that the critical period begins several months after birth and extends to approximately 3 years of age. Comparison of functional results from surgical correction of early-onset esotropia with age at correction shows that no patients surgically corrected after the age of 2 years exhibited stereopsis, whereas a high proportion of those who were corrected before that age have demonstrable stereopsis.55,56 It has been suggested both directly and indirectly in this discussion that the presence and level of stereoacuity may depend on certain innate factors beyond those measured by the amount of interocular transfer exhibited and the presumed proportion of binocularly driven cortical neurons.
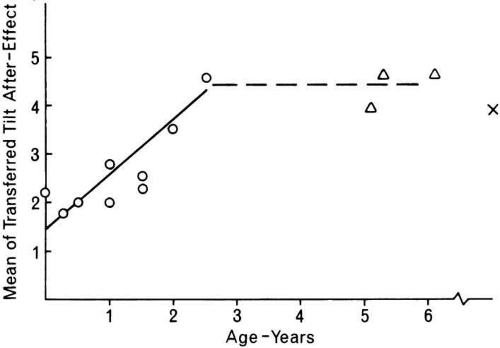 Fig. 5 Interocular transfer of aftereffects as a function of age at onset of strabismus. Strabismus after age 3 caused no reduction in interocular transfer relative to adult levels (x). (Hohmann A, Creutzfeldt OD: Squint and the development of binocularity in humans. Nature 254:613, 1975) |
STRABISMIC AMBLYOPIA
Amblyopia, a loss of spatial resolution without overt retinal pathology, occurs in two major forms differing in cause and expression.57 One is strabismic amblyopia, which is the type associated with eye misalignment. Functional competition between the two eyes seems to be a major factor in this condition. Differences between the eyes in image focus (anisometropia) or image magnification (aniseikonia), or partial occlusion of the image in one eye also will lead to a preference for one eye, so that the other eye develops a strabismic misalignment and then amblyopia.57,58,59 Even though the retinal images may be clear in each eye, one eye may be used for fixation more than the other (e.g., because it has better motor performance), with a resulting amblyopia in the less dominant eye. However, if each eye is used a portion of the time, as in alternating esotropia, asymmetric amblyopia is unlikely to develop. Contrary to the conventional view, however, one report60 shows that most alternating esotropes exhibit mild bilateral amblyopia, an equal reduction in acuity for both eyes relative to normal, in addition to the expected loss of stereopsis and binocular function. Finally, profound deprivation amblyopia in one eye may lead to strabismus of that eye, for example, in congenital cataract.58 Thus, all mixtures of deprivation and strabismic amblyopia occur.
Amblyopia in strabismus is more often associated with eccentricity of fixation than is mild deprivation amblyopia. In approximately 50% of comitant esotropia cases, the position of the retina used for monocular fixation by the amblyopic eye corresponds with its angle of strabismus.60
Strabismic amblyopia is frequently seen in infants as young as 4 or 5 months of age; such an esotropic eye (misaligned toward the nose) may not even be able to fix a target. This fact supports the suggestion54 that the “sensitive period” for strabismic amblyopia in humans has its beginning before 5 months. Conversely, amblyopia from a strabismus beginning after the age of 5 or 6 years is rare.
DEPRIVATION AMBLYOPIA
The second major category is deprivation amblyopia, which occurs when the image in one eye or both eyes is not normally clear. This condition may be from refractive error (astigmatism, aphakia), from interference with clear optics (corneal scar, cataract), or from glare degrading the retinal image (aniridia). The visual consequences depend on the severity of deprivation, difference of deprivation between the two eyes, age at onset, and duration before treatment.
Profound defects in retinal imagery from birth (e.g., dense cataract of the lens) affect the visual system so strongly that removal of the cataractous lens after 2 years of age, even given a clear image and occlusion treatment of the good eye (if unilateral), is insufficient to restore useful vision above approximately 20/200 (0.1).
Blur, if not too severe, may leave the visual acuity open to marked improvement through optical correction, even though the eye was deprived of clear vision during much of the development period. As an example, bilateral hyperopia of +8 diopters (that was present since infancy), when first corrected by glasses at age 5 or 6, typically gives a best corrected vision of 20/100 (0.2). However, development of the vision over 2 to 3 years with corrective glasses results in a visual acuity of almost 20/20 (1.0). Persistence of some amblyopia is found for the more hyperopic meridian in hyperopic astigmatism.62
Besides the severity of blur, the difference in image clarity of the two eyes determines the profoundness of amblyopia. It is a common clinical occurrence to see cataracts develop or progress at age 3 or 4 years, resulting in vision of 20/200 or less. When vision is equal in both eyes, amblyopia does not usually occur and 20/20 vision is often restored by surgery. On the contrary, a unilateral cataract of the same severity at such an age is a visual disaster unless rapidly treated, resulting in profound amblyopia.63
If sufficiently severe, a deprivation in infancy cannot be reversed even after a few months of age. Although this susceptibility progressively diminishes, untreated traumatic cataracts without other injury to the eye, acquired even after the age of 8, can still result in permanent amblyopia. However, after approximately the age of 10, amblyopia can no longer be produced even by the “total occlusion” of a cataract. The period of amblyopia sensitivity is over.64
An interesting prediction derived from the binocular competition hypothesis of the effects of monocular deprivation is that form deprivation amblyopia should be minimized by the previous occurrence of misalignment between the eyes,65 because an early strabismus with alternating fixation generates a strong independence between the cortical representations of the two eyes. The predicted sparing of the deprived eye from profound amblyopic loses has been found in cats65 and monkeys.66 Paradoxically, therefore, previously strabismus can protect the visual system from the severe effects of monocular deprivation.29
Reports67,68,69 suggest that deprivation amblyopia can be improved at least transiently in human adults and children by the oral administration of levodopa. Visual acuity, contrast sensitivities, and VEP amplitudes were significantly improved and the size of amblyopic suppression scotoma reduced by administration of this neurotransmitter to the amblyopic patients.
A final aspect of such visual deprivation is a loss of binocular function, even without an amblyopic acuity loss. Even in children as old as 10 to 18 years, the occurrence of cataract by injury may sometimes cause a loss of binocular fusion with resulting diplopia if removal of the cataract is delayed up to 3 to 4 years. Thus, the period of susceptibility to binocular disruption may persist through adolescence, a much longer period than for strabismic amblyopia.
STEREOSCOPIC VISION
BINOCULAR DISPARITY
Stereopsis was discussed briefly on p. 3, but will now be considered in greater detail. Stereoscopic vision may be defined as the ability to see depth in the third dimension of visual space (i.e., distance from the observer) on the basis of relative image positions of the two retinas. The fact that the two eyes are horizontally separated in space implies that, in general, they will receive slightly disparate views of all objects located at a distance different from the convergence distance. The field of horizontal binocular disparities, combined with information as to the fixation directions of the two eyes (convergence), provides a precise quantitative description of the distance of the objects in the field.
Considering the situation close to the line of sight for simplicity, if both eyes fixate a point object at a given distance (bifoveal fixation), the image of that object obviously will fall on the fovea in each eye (point F in Fig. 3). There then will be no binocular disparity between the two monocular images. If a second object represented by the arrowhead is now presented at a different distance from the first, a difference in depth may be signaled in two ways: binocular disparity and convergence angle
If we consider the retinal projections of the head and tail of the arrow, there is now a binocular disparity between the projections to the two eyes (see Fig. 3). This disparity can be interpreted neurally as a spatial depth relative to the point of convergence. When the object is in front of the point of convergence, the lines of sight of the monocular images cross before the convergence distance, giving rise to a crossed disparity. If, however, the object were moved away from the observer, the lines of sight would not cross before the convergence distance, and the disparity would be described as uncrossed.
Note that it is a mistake to regard the nasal or temporal retina as being associated with specific depth information, as has sometimes occurred. A nasal shift in crossed disparity and a temporal shift for uncrossed disparity do not mean that the images fall on the nasal and temporal retina, respectively, except when the images are at or close to the fovea. Thus, if the near object is viewed at 5 degrees eccentricity to the left, both images will fall to the right of the forca for all disparities (up to 5 degrees), whether the disparity is near, zero, or far.
A second result of moving the object closer is that the eyes may converge to the new vergence angle so as to reacquire bifoveal fixation on the object. The change in vergence angle provides the cue to the new distance of the object, because the binocular disparity is again zero. Thus, vergence angle is a cue to depth that is distinct from, and interacts with, retinal disparity. It may be sensed variously by muscular feedback, by eye movement control signals, or by changes in the vertical disparity fields between the eyes.70
PSYCHOPHYSICAL STUDIES
PRECISION OF STEREOSCOPIC LOCALIZATION
Under normal conditions, most observers with no ocular abnormalities can discriminate a depth difference between two images with a relative disparity as small as 10 arc-seconds (0.0028 degrees). The best values reported in the literature are obtained using a method of constant stimuli, in which the observer is presented with a series of disparities between a test and comparison rod stimuli and asked to discriminate whether the test rod is nearer or farther than the comparison rods.71 Whether monocular information is present or is eliminated, the best observers achieve a 75% discrimination level close to 2 arc-seconds (0.00056 degrees).
This value appears to be one of the finest spatial discriminations of which the human visual system is capable. It represents a truly amazing accomplishment, particularly considering that the resolution limit for dark lines is more than 10-times larger at the intensities used in the stereoscopic discrimination tasks and also that the discrimination necessitates comparing between two retinas in continuous motion because of eye movements.72 To illustrate the refinement of this discrimination, it can be converted into real distances for the near and far limits of vision. For the closest fixation of 10 inches away, the best stereoscopic threshold corresponds to the appreciation of a depth of one-thousandth of an inch (25 μm). For comparison, this performance is finer than the size of a typical human ovum (100 μm) or the cell body of a typical neuron (50 μm). Conversely, when looking at the horizon, one has stereoscopic discrimination that objects 2 miles away are being nearer than the horizon. This limit allows, for example, stereoscopic discrimination of depth in some types of cloud formations. Contrary to common statements that stereopsis is useful only in the near field, these limits provide useful stereopsis over an extensive range of environmental conditions.
SPATIAL LIMITS OF STEREOPSIS
Although stereoacuity is excellent at the fovea, it diminishes with a steepening function as the stimulus is moved into the periphery, so that stereopsis becomes very poor beyond approximately 20 degrees eccentricity.73
Just as binocular disparity can be too small to elicit a perception of depth, disparity can be increased until it is too large for depth perception. As pointed out earlier, depth continues to be perceived well beyond the range of binocular fusion. The perceived depth with disparity is shown as a function of eccentricity in Figure 6. The stimuli were perceived as fused only up to just before the peak of each function. Because it has a logarithmic axis, this graph makes the important point that there is larger region of stereopsis for which the stimuli are diplopic than there is when they are fused. In fact, the greatest perceived depth for each stimulus size is found beyond the region of fusion.
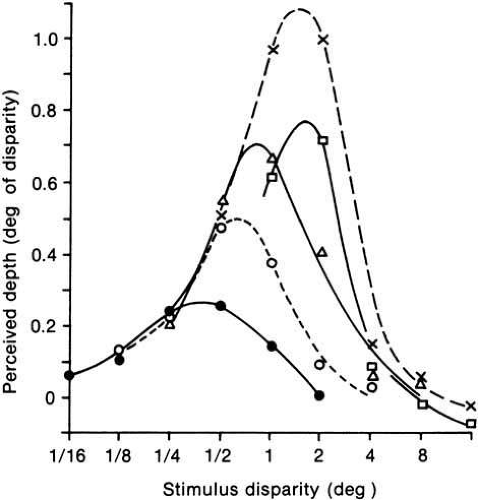 Fig. 6 Relative perceived depth versus stimulus disparity averaged for three observers. Near (convergent) disparities only. Each curve represents a separate bar width: dots, 0.5 degrees; open circles, 0.1 degrees; triangles, 0.2 degrees; crosses, 0.4 degrees; squares, 0.8 degrees. Bar height was fixed at 0.5 degrees. Stimuli were perceived as fused only on the rising portion of the curves. (Richards W, Kay MG: Local versus global stereopsis: Two mechanisms? Vision Res 12:437, 1972) |
Ogle73 refers to these regions as “quantitative” (for fused) and “qualitative” (for diplopic) stereopsis, but his terms are inaccurate because the perceived depth may be quantified in both regions74 (see Fig. 6). It therefore seems more appropriate to designate the two regions as “fused” and “diplopic” stereopsis, thereby avoiding pejorative implications.
Another property that is somewhat related to these two regions is that perceived depth increases linearly with binocular disparity in most of the fused region, whereas it falls with some nonlinear function through the diplopic region. However, the transition from a linear to nonlinear function does not occur at exactly the same disparity as the transition from fusion to diplopia, so the correspondence between the two divisions is not precise.
A key aspect of Figure 6 is the change in the depth magnitude function for different widths of the text bar.74 The maximum depth occurs at disparities varying from 0.5 degrees to approximately 2 degrees; a disparity that can produce no depth impression at all for a small stimulus (e.g., 1 degree) may be optimal for depth perception in larger test objects (see Fig. 6). This result suggests that tests for stereoscopic vision should involve stimuli with a good range of sizes and disparities to be sure of the capabilities of an abnormal visual system.
Another way of demonstrating the effect of stimulus size on stereoscopic ability was described by Tyler.75,76 The maximum disparities for which depth was perceived were inversely proportional to vertical length of the segments over a range from 0.05 degrees to 5 degrees. This behavior was termed disparity scaling of the upper depth limit. Such disparity scaling reduces the complexity in processing the three-dimensional image while retaining the full range of sensitivity to disparities from seconds to degrees. Thus it is possible to appreciate many aspects of a three-dimensional scene without requiring the order of magnitude of more neurons that would be required for full depth processing to the same resolution as for each eye alone.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


