Aqueous Humor Dynamics
J. Cameron Millar
B’Ann True Gabelt
Paul L. Kaufman
In the healthy eye, flow of aqueous humor against resistance generates an intraocular pressure of about 15 mmHg, which is necessary for the proper shape and optical properties of the globe.1 The circulating aqueous humor nourishes the cornea and lens, structures which must be transparent and therefore devoid of blood vessels;1 the aqueous also provides a transparent and colorless medium of refractive index 1.3333 between them,2 and thus is an important component of the eye’s optical system.1 The basic anatomy of the primate anterior ocular segment and the normal pathways of aqueous humor flow are shown schematically in Figures 1 and 2.3 Aqueous humor is secreted by the ciliary epithelium lining the ciliary processes (by active ionic transport across the ciliary epithelium and hydrostatic and osmotic gradients between the posterior chamber and the ciliary process vasculature and stroma). It enters the posterior chamber and then flows around the lens and through the pupil into the anterior chamber, from which it leaves the eye by passive bulk flow through two pathways at the anterior chamber angle: (1) through the trabecular meshwork (TM), across the inner wall of Schlemm’s canal and into its lumen, and then into collector channels, aqueous veins, and the general episcleral venous circulation—the trabecular or conventional route; and (2) across the iris root, uveal meshwork, and anterior face of the ciliary muscle, through the connective tissue between the muscle bundles, and the suprachoroidal space, and then out through the sclera—the uveoscleral, posterior, or unconventional route. In certain monkey species, total aqueous drainage is relatively evenly divided between the two pathways.4,5,6,7 In the normal human eye, the importance of the uveoscleral pathway has not been well determined. In elderly eyes with posterior-segment tumors, it accounts for about 10% of total aqueous humor drainage, but in young individuals it may account for more than 30%.8,9,10 There is no significant net fluid movement across the cornea, iris vasculature, or vitreoretinal interface, although ion fluxes exist.11,12
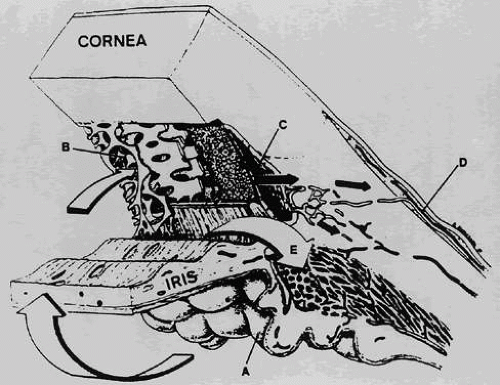 Fig. 1. Cross-section through the anterior segment of the eye illustrating the chamber angle. Aqueous humor is formed by active secretion from the ciliary processes (A). Drainage occurs through the outflow pathways, principally from the trabecular meshwork (B) and Schlemm’s canal (C) into the aqueous veins (D). A smaller proportion of the aqueous humor makes its way directly into the ciliary body (uveoscleral pathway) and is drained by way of the ciliary muscle, the suprachoroidal space, and the sclera (E). (Adapted from Karnezis TA, Murphy MB: Dopamine receptors and intraocular pressure. Trends Pharmacol Sci 9:389, 1988) |
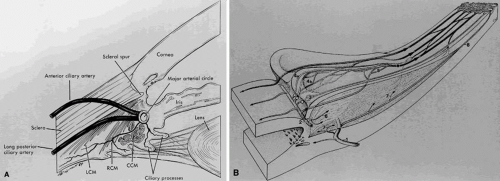 Fig. 2. A. Blood supply to the ciliary processes. LCM, longitudinal ciliary muscle; RCM, radial ciliary muscle; CCM, circular ciliary muscle. B. Vascular architecture in the human ciliary body. (1) Perforating branches of the anterior ciliary arteries; (2) major arterial circle of iris; (3) first vascular territory; the second vascular territory is depicted in (4a) marginal route, and (4b) capillary network, in the center of this territory; (5) third vascular territory; (6 and 7) arterioles to the ciliary muscle; (8) recurrent choroidal arteries. Light circles, terminal arterioles; dark circle efferent venous segment. (A, Caprioli AJ: The ciliary epithelia and aqueous humor. In Hart M [ed]: Adler’s Physiology of the Eye, p 228. 9th ed. St. Louis: Mosby, 1992; and B, Rohen JW, Funk RH: Vasculature of the anterior eye segment. Prog Ret Eye Res 13:653, 1994) |
Because of autonomic innervation and the receptors of the relevant structures, adrenergic and cholinergic mechanisms play major roles in aqueous humor formation and drainage in terms of both normal physiology and glaucoma therapeutics13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32; evidence for other mechanisms, including serotonergic,33,34,35,36 dopaminergic,37,38,39,40,41,42,43,44,45,46 adenosinergic,47,48,49,50,51,52 and prostaglandinergic mechanisms 6,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67 is growing.
AQUEOUS HUMOR FORMATION AND COMPOSITION
PHYSIOLOGY
Until the early twentieth century, aqueous humor was regarded as a stagnant fluid.68 Since that time, however, it has been shown to be continuously formed and drained,69 and the associated anatomic drainage portals (Schlemm’s canal, collector channels, aqueous veins, and ciliary muscle interstices) have been described.70,71,72,73
Three physiologic processes contribute to the formation and chemical composition of the aqueous humor1: diffusion, ultrafiltration (and related dialysis), and active secretion. The first two processes are passive and therefore require no active cellular participation. Diffusion of solutes across cell membranes occurs down a concentration gradient, and substances with high lipid solubility coefficients that can easily penetrate biological membranes move readily in this way. Ultrafiltration is the term used to describe the bulk flow of blood plasma across the fenestrated ciliary capillary endothelia into the ciliary stroma; it can be increased by augmentation of the hydrostatic driving force (see Fig. 2). These processes (diffusion and ultrafiltration) are responsible for the formation of the “reservoir” of the plasma ultrafiltrate in the stroma, from which the posterior chamber aqueous is derived through active secretion across the ciliary epithelium. Active secretion requires energy, normally provided by the hydrolysis of adenosine triphosphate (ATP). The energy is used to secrete substances against a concentration gradient. Energy-dependent active transport of sodium into the posterior chamber by the nonpigmented ciliary epithelium (NPE) (see Fig. 2) results in water movement from the stromal pool into the posterior chamber. Although controversy has surrounded the relative quantitative roles of ultrafiltration and active secretion, it seems fairly certain that under normal conditions active secretion accounts for perhaps 80% to 90% of total aqueous humor formation.1,11,74,75,76,77,78,79,80,81 The observation that moderate alterations in systemic blood pressure and ciliary process blood flow have little effect on aqueous formation rate supports this notion.11,74,82 Moreover, Bill noted that the hydrostatic and oncotic forces that exist across the ciliary epithelium—posterior aqueous interface favor resorption, not secretion, of aqueous humor.74 Active secretion is essentially pressure-insensitive at near-physiologic intraocular pressure (IOP). However, the ultrafiltration component of aqueous humor formation is sensitive to changes in IOP, decreasing with increasing IOP. This phenomenon is quantifiable and is termed facility of inflow or pseudofacility (Cps)—the latter because a pressure-induced decrease in inflow appears as an increase in outflow when techniques such as tonography and constant-pressure perfusion are used to measure outflow facility.1,83,84,85,86,87,88,89 Cps quantities in monkeys87,90 and humans91 are approximately 0.02 μl and 0.06 μl × min-1 × mmHg-1, respectively, although the latter may be an overestimate.
In most mammalian species, the turnover constant of the anterior chamber aqueous humor is approximately 0.01 × min-1, that is, the rate of aqueous humor formation and drainage is about 1% of the anterior chamber volume per minute.76,92,93,94,95,96 This is true also in the normal human eye, in which the aqueous formation rate is approximately 2.0 to 2.5 μl × min-1.1,8,97,98,99,100,101
BIOCHEMISTRY
The active process of aqueous secretion is mediated through selective transport of certain ions and substances across the basolateral membrane of the NPE against a concentration gradient. Two enzymes abundantly present in the NPE are intimately involved in this process: sodium-potassium-activated adenosine triphosphatase (Na+ -K+ -ATPase) and carbonic anhydrase (CA).78,106,107,108,109,110,111,112,113,114,115,116,117 Na+ -K+ -ATPase is found predominantly bound to the plasma membrane of the basolateral infoldings of the NPE.78,108,109,110,111,113,114116 The enzyme provides the energy for the metabolic pump, which transports sodium into the posterior chamber, by catalyzing the reaction ATP → ADP + Pi + energy,118,119 where ATP = adenosine triphosphate, ADP = adenosine diphosphate, and Pi = inorganic phosphate.
As a result of active transport, aqueous humor in humans exhibits greater levels of ascorbate, some amino acids, and certain ions such as Cl-.99,101 There is also a passive transporter for HCO3-.1,120 A summary of the biochemistry of aqueous secretion is shown in Figure 3.
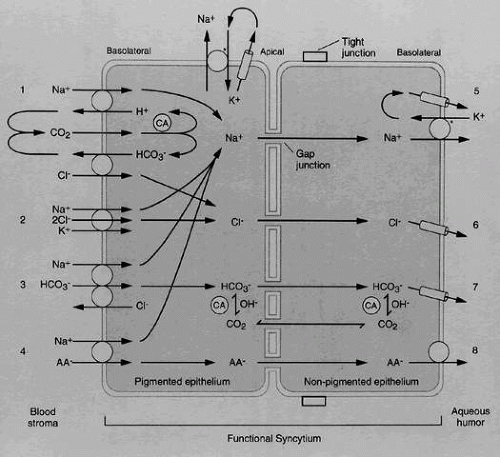 Fig. 3. Diagram of possible secretory pathways in the ciliary processes. AA, ascorbic acid; CA, carbonic anhydrases. (Adapted from Wiederholt M, Helbig, H, Korbmacher C: Ion transport across the ciliary epithelium: Lessons from cultured cells and proposed role of the carbonic anhydrase. In Botné F, Gross G (eds): Carbonic Anhydrase, p 232. Basel: Verlag-Chemie, 1991) |
Inhibition of the ciliary process Na+ -K+ -ATPase by cardiac glycosides (e.g., ouabain) or vanadate (VO3-, VO43-) significantly reduces the rate of aqueous humor formation and consequently IOP in experimental animals108,113,119,121,122,123,124 and humans.123 Cardiac glycosides appear to act at the extracellular aspect of the membrane-bound enzyme, but vanadate acts at the cytoplasmic surface.124,125 Cardiac glycosides given topically are ineffective as ocular hypotensives and may cause corneal edema by interfering with the Na+ -K+ -ATPase—dependent sodium pump in the corneal endothelium.121 Intravitreal and systemic administration are effective but carry unacceptable ocular and cardiovascular risks, respectively.121,126 Vanadate is effective topically in both rabbits and monkeys, apparently without producing acute corneal edema.119,122,123
CA is abundantly present in erythrocytes, renal tubules, and the basal and lateral membranes and cytoplasm of the pigmented epithelium and NPE of the ciliary processes.80,127,128,129,130,131,132 CA catalyzes reaction I of the sequence
CO2 + H2O →I H2CO3 →II H+ + HCO3–
Reaction II is a spontaneous, virtually instantaneous ionic dissociation.118,131 The actual sequence may be far more complex, involving energy-dependent separation of H+ and OH- at membrane boundaries within the NPE cell, and formation of HCO3– by CA-catalyzed association of OH- with CO2.80,133 The reaction sequence shown above provides the HCO3-, which is essential for the active secretion of aqueous humor. Although the exact roles of CA and HCO3– are still debated,76,78,80,120,133,134,135,136 it has been demonstrated that inhibition of the production of HCO3– also leads to an inhibition of the active transport of Na+ across the NPE into the first-formed aqueous, thereby reducing active aqueous humor formation (AHF). Several hypotheses explain the relationship between reduction in NPE intracellular HCO3– and inhibition of Na+ transport,1 including: (1) inhibition of CA causes a decrease in HCO3– available for transport with Na+ from the cytosol of the NPE to the aqueous, which is required to maintain electroneutrality; (2) reduction in intracellular pH inhibits Na+ -K+ -ATPase, and (3) decreased availability of H+ produced by reaction II decreases H+ /Na+ exchange and reduces the availability of intracellular Na+ for transport into the intercellular channel. In addition, inhibition of renal and erythrocyte CA leads to a systemic acidosis that promotes inhibition of AHF.137
CA inhibitors (e.g., acetazolamide,138 methoxazolamide, ethoxzolamide, dichlorphenamide,129,139 aminozolamide,140 trifluormethazolamide141) given systemically can reduce secretion by as much as 50%127,128,129,132,139,140,141,142,143,144,145,146,147,148 and have been in use for clinical glaucoma therapy for more than 40 years.135,136,139,149 With low doses of certain CA inhibitors (e.g., methazolamide), it is possible to inhibit the ocular enzyme without affecting the renal and erythrocyte enzymes, thus producing submaximal secretory suppression without systemic acidosis.80,129,139,150,151 It was once thought that the drug concentration at the ciliary epithelium required to produce the almost continuous, total ciliary CA inhibition necessary to achieve adequate and sustained reduction in AHF (over 99% of the ciliary enzyme must be inhibited to achieve significant secretory suppression) might never be attainable through the eye-drop route,80,129,136,139,151, but the topically effective CA inhibitor dorzolamide,146,147,152,153,154,155 which may achieve nearly the same reduction in IOP as the earlier CA inhibitors but without their systemic side effects, is currently available.137
AQUEOUS HUMOR COMPOSITION
The composition of aqueous humor differs from that of plasma as a result of two important physiological characteristics of the anterior segment: a mechanical epithelial/endothelial blood-aqueous barrier, and active transport of various organic and inorganic substances by the ciliary epithelium. The greatest differences are the low protein and high ascorbate concentrations in the aqueous relative to plasma (about 200 times less and 20 times greater, respectively).156,157,158,159,160,161,162 The high ascorbate concentration may help protect the anterior ocular structures from ultraviolet light-induced oxidative damage. When the aqueous protein concentration rises significantly above its normal level of approximately 20 mg/100 ml,76 as in uveitis, the resultant light scattering (Tyndall effect) makes visible the slit-lamp beam as it traverses the anterior chamber (a phenomenon known as “flare”). Lactate also is normally in excess in the aqueous, presumably as a result of glycolytic activity of the lens, cornea, and other ocular structures.76,143 Other compounds or ions in excess in the aqueous relative to the plasma are Cl- and certain amino acids.101
BLOOD-AQUEOUS BARRIER
The blood-aqueous barrier (BAB)is a functional concept, rather than a discrete structure, invoked to explain the degree to which various solutes are relatively restricted in travel from the ocular vasculature into the aqueous humor. The capillaries of the ciliary processes and choroid are fenestrated, but the interdigitating surfaces of the retinal pigment epithelia and the ciliary process NPE respectively are joined to each other by tight junctions (zonulae occludens) and constitute an effective barrier to intermediate- and high-molecular weight substances, such as proteins.163,164,165,166,167,168,169,170,171,172,173 The endothelia of the inner wall of Schlemm’s canal are similarly joined,166,174 preventing retrograde movement of solutes and fluid from the canal lumen into the TM and anterior chamber. The iris and retina have no similar epithelium between their vasculature and the ocular fluids, but their capillaries are of the nonfenestrated, impermeable type.11,76,101,165,168 For present purposes, one may say that the BAB comprises the tight junctions of the ciliary process NPE, the inner wall endothelium of Schlemm’s canal, the iris vasculature, and the outward-directed active transport systems of the ciliary processes. A more universal concept of the BAB must explain the movement of smaller molecules, lipid-soluble substances, and water into the eye.76
With disease-, drug-, or trauma-induced breakdown of the BAB (Table 1), plasma components enter the aqueous humor. Net fluid movement from blood to aqueous increases, but so does its IOP-dependence (Cps).175 Total facility, as measured by IOP-altering techniques, cannot distinguish Cps from total outflow facility (Ctot) and therefore erroneously records the Cps component as increased Ctot (hence the term, ‘pseudofacility’) and underestimates the extent to which the outflow pathways have been compromised by the insult. Under these circumstances, increased Cps provides some protection against a precipitous rise in IOP; as IOP rises, aqueous inflow by ultrafiltration is partly suppressed, blunting (but not completely suppressing)88,176 further IOP elevation. Additionally, the inflammatory process that occurs during BAB breakdown leads to a reduction in active secretion of aqueous humor, possibly through interference with active transport mechanisms.177 This in turn may actually produce ocular hypotony, despite compromised outflow pathways (because of plasma protein blockage of the TM). Prostaglandin release during inflammation may contribute to the hypotony by increasing aqueous outflow through the uveoscleral route.178,179 When the noxious stimulus is removed, however, the ciliary body may recover before the TM, and the resulting normalization of AHF rate in the face of still-compromised outflow pathways leads to elevated IOP, as seen from the modified Goldmann equation IOP = [(F – U)/Ctrab] + Pe, where F = aqueous humor flow, Ctrab = facility of outflow from the anterior chamber through the trabecular meshwork and Schlemm’s canal, IOP = intraocular pressure, Pe = episcleral venous pressure (the pressure against which fluid leaving the anterior chamber through the trabecular-canalicular route must drain), and U = uveoscleral outflow.180
TABLE 45-1. Factors Interrupting the Blood-Aqueous Barrier | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
ACTIVE TRANSPORT
The ciliary processes possess the ability to actively transport (or exclude) a variety of organic and inorganic compounds and ions from the eye, that is, to move them from the aqueous or vitreous to the blood against a concentration gradient. Para-aminohippurate (PAH), diodrast, and penicillin are examples of large anions that are actively transported out of the eye. These systems are similar to those in the renal tubules and satisfy all the criteria for active transport, including saturability, energy and temperature dependence, Michaelis-Menten kinetics, and inhibition by ouabain and probenecid among others.101,181,182,183,184,185,186,187,188,189,190,191,192 In addition, another system actively excretes injected iodide from the aqueous, resembling iodide transport in the thyroid and salivary glands.187 The physiologic role of these outward-directed systems is unknown. With the discovery that prostaglandins may be actively transported out of the eye,193,194 some workers have suggested that such outward-directed mechanisms may rid the eye of biologically active substances that are no longer needed or may even be detrimental.186,188,195,196,197 Other outwardly-directed ion-uptake mechanisms are present in the eye. The anterior uvea of the rabbit eye, for example, accumulates the anions cholate, glycocholate, deoxycholate, chenodeoxycholate, iodipamide, and o-iodohippurate.181,185 At least one outwardly-directed cationic pump also has been reported; iris-ciliary body (ICB) preparations accumulate the cation emepronium,198 although one report199 questioned whether any other cations are actively eliminated from the eye.
Bárány,183 making an analogy to the ion pump located at the renal peritubular cell border adjacent to the blood, which pumps simple cations from the blood to the kidney tubule, investigated whether there are any inwardly-directed complex cation pump systems from blood to aqueous, but concluded that such mechanisms probably do not exist, at least in the rabbit. All of these transport systems (inward- and outward-directed) are thought to be located in the NPE.76
PHARMACOLOGY AND REGULATION
Sympathetic and parasympathetic nerve terminals are present in the ciliary body200,201,202,203,204 and arise from branches of the long and short posterior ciliary nerves. These nerve fibers are of both myelinated and nonmyelinated variety. Parasympathetic fibers originate in the Edinger-Westphal nucleus of the third cranial nerve, run with the inferior division of this nerve in the orbit, and synapse in the ciliary ganglion.205 Sympathetic fibers synapse in the superior cervical ganglion and are distributed to the muscles and blood vessels of the ciliary body. Numerous unmyelinated nerve fibers surround the stromal vessels of the ciliary processes; these are most likely noradrenergic and subserve vasomotion.76 Sensory fibers arise from the ophthalmic division of the trigeminal nerve and enter the ciliary body, but their distribution and function have not been studied thoroughly. No innervation of the ciliary epithelium has been found anatomically,166 but stimulation of the ciliary ganglion leads to an increase in AHF in the enucleated arterially perfused cat eye,206 suggesting that neurotransmitters released in the ciliary stroma might diffuse toward the epithelia.
Cholinergic Mechanisms
The effects of cholinergic drugs on AHF and composition and on the BAB are unclear. In general, cholinergic drugs cause vasodilation207,208,209 in the anterior segment,210,211,212,213 resulting in increased blood flow to the choroid, iris, ciliary processes, and ciliary muscle.210,211,214 However, cholinergic drugs also may promote vasoconstriction—in rat coronary arteries208 and rat outer descending vasa recta perfused in vitro,215 for example, and in the rabbit eye.216 These responses are mediated by muscarinic receptors in the anterior uveal arterioles,210 perhaps associated with facial parasympathetic nerve terminals.217,218 Congestion in the iris and ciliary body is a well-recognized clinical side effect of topical cholinomimetics, especially the anticholinesterases.219 The presence of flare and cells in the aqueous humor seen with biomicroscopy indicates that these agents also can cause breakdown of the BAB and perhaps frank inflammation.219 Pilocarpine increases BAB permeability to iodide220 and inulin.221 Cholinergic drug-induced vasodilation may cause a loss of tight junctions in anterior uveal blood vessels, perhaps contributing to BAB breakdown.222 Cholinergic drugs may alter the aqueous humor concentration of inorganic ions223 and the movement of certain amino acids from the blood into the aqueous humor and may also influence the outward-directed transport systems of the ciliary processes.224,225
Under certain conditions, pilocarpine may increase Cps.84,91 With a variety of species, conditions, and experimental techniques, cholinergic agents or parasympathetic nerve stimulation has been reported to increase, decrease, or not alter the AHF rate and to increase slightly the episcleral venous pressure.91,206,225,226,227,228,229,230,231,232,233,234,235,236,237,238,239 These apparently confusing results may indicate that cholinergic drug effects on these parameters are extremely dependent on species- and technique-related factors and on the ambient neurovascular milieu. In any event, the effects on the rate of AHF and episcleral venous pressure are surely minor in most instances, and not responsible for the drug-induced decrease in IOP that forms the basis of pilocarpine’s therapeutic efficacy in chronic glaucoma; the latter resides in its ability to decrease outflow resistance through its effect on the ciliary muscle.
Adrenergic Mechanisms
The precise role and receptor specificity of adrenergic mechanisms in regulating the rate of AHF are unclear. At one time it was generally believed that long-term topical administration of epinephrine, a combined α1-, β1-, β2-adrenergic agonist, would decrease the rate of AHF.240 This effect was thought to be mediated by β-adrenergic receptors in the NPE, through activation of a membrane adenylate cyclase,115,241,242,243 although the consequent biochemical events were (and still are) unknown. In support of these observations, activation of ocular adenylate cyclase through its Gs protein in the rabbit by close arterial infusion of cholera toxin decreases AHF and lowers IOP.241 Further, forskolin, a naturally occurring diterpene derivative of the coleus plant (Coleus forskohlii) that directly and irreversibly activates intracellular adenylate cyclase in some244,245,246,247,248,249 but not all250 studies, decreases the rate of AHF when given topically or intravitreally.
Although vascular phenomena could theoretically be involved in the secretion and pressure responses, these observations are consistent with the primary epithelial action of these drugs. The relationship between ocular vascular events and IOP modulation, if any, has yet to be established.103,251,252,253,254,255,256
Recent fluorophotometric studies have shown that short-term topical administration of epinephrine increases AHF8,13,29; studies with other adrenergic agonists, including salbutamol,257 isoproterenol (isoprenaline),258 and terbutaline,100 have supported this finding and are consistent with many studies showing that β-adrenergic antagonists unequivocally decrease AHF.14,18,19,20,21,23,24,25,26,27,29,31,32,97
The ocular hypotensive action of β-antagonists has led to their becoming mainstays of clinical glaucoma therapy; they include nonselective β1, β2 antagonists timolol,20,21,29,97 levobunolol259, and metipranolol;24 the nonselective β1, β2 partial agonist carteolol14; and the relatively selective β1 antagonist betaxolol.31 Adrenergic receptors in the ciliary epithelium are of the β2 subtype,260,261,262,263 but antagonists that are relatively selective for β1 receptors (e.g., betaxolol) are effective (although less potent and efficacious)264,265 in suppressing AHF.31,266,267,268,269 However, the apparent β1 efficacy may be related to a sufficiently high concentration reaching the ciliary epithelium so that nonselective blockade of β2 receptors may occur.
Whether β antagonists suppress AHF through their effect on ciliary epithelial β receptors has been questioned.119,233,241,244,245,246,248,249,250,270,271,272,273,274,275,276,277,278,279,280 There is evidence that classical β-blockade may not be involved, and that other receptor types such as 5-HT1A, may be relevant (see later).33,35,36 Furthermore, inhibition of Na/K/2Cl cotransport by β2 receptor antagonists can eliminate the increase in AHF stimulated by epinephrine and isoproterenol.281,282
AHF is reduced by nearly 50% during sleep,76,101 which is comparable to the daytime reduction induced by β-adrenergic antagonists, or to the daytime or nighttime reduction induced by the CA inhibitors.124 β-antagonists produce little additional decrease in AHF during sleep98 or in pentobarbital-anesthetized monkeys.283 Because sympathetic tone is reduced during both sleep and barbiturate anesthesia,284 these findings provide further indirect evidence that β-adrenergic tone/stimulation enhances AHF but β-adrenergic blockade decreases it.
Topically applied α1-adrenergic agonists and antagonists appear to have little effect on fluorophotometrically-determined AHF in the normal intact human eye,285,286 although these compounds do reduce AHF in rabbits.287,288,289,290,291 However, in eyes under β-adrenergic blockade, topical epinephrine acutely produces a small decrease, rather than increase, in AHF, perhaps indicating a weak α-adrenergic influence, possibly but not necessarily mediated by local vasoconstriction.292,293 Clonidine, which has both α1-antagonist and α2-agonist properties, decreases AHF and ocular blood flow.294,295,296,297 Therefore epinephrine may have a dual effect on AHF: stimulation through β-adrenoreceptors, and inhibition through α2-adrenoreceptors.298,299,300,301 α2-Adrenergic agonists such as apraclonidine HCl (AP) and brimonidine tartrate (BR) are powerful ocular hypotensive agents when applied topically. Both AP and BR are believed to lower IOP primarily by decreasing AHF.302
There are many other ways in which AHF can be reduced pharmacologically (Table 2). Effective compounds include: the guanylate cyclase activators, atrial natriuretic factor (ANF),303 the nitrovasodilators, sodium nitroprusside,304,305 sodium azide,304 and nitroglycerin.306 8-Bromo cyclic GMP also reduces the AHF rate by 15% to 20% in the monkey.248 ANF injected intravitreally reduced IOP and AHF in rabbits and monkeys.307,308 The calcium channel antagonists verapamil and nifedipine have been reported to reduce AHF in rabbits,309 although topical diltiazem and verapamil have been reported to increase AHF in human volunteers.310 The serotonergic antagonist ketanserin reduces the AHF rate in rabbits, cats, and monkeys.311,312 The endogenous agonist, serotonin (5-HT) may also reduce AHF; topical application results in decreased AHF in the rabbit,312 but intracameral injection in the same species increases AHF.312 Serotonergic receptors of a 5-HT1A-like subtype have been reported to exist in the ICB of rabbits and humans.33,36 It has been suggested that these receptors may be antagonized by timolol and other β-blockers. However, the precise nature of the putative 5-HT1A-like receptor subtype in the ciliary epithelium is still in question. Angiotensin converting enzyme (ACE) is present in human and rabbit aqueous humor24,25,26 and also in the feline ciliary body.27 Several ACE inhibitors, when topically applied to rabbit or human eyes, reduce secretion. H1-antihistamines (such as antazoline and pyrilamine) decrease AHF in rabbits,313 although the effect may be unrelated to histamine receptor binding. Δ9-tetrahydrocannabinol (a component of marijuana) may reduce AHF in humans when injected intravenously or inhaled (as in marijuana smoking).314 Metabolic inhibitors such as dinitrophenol (DNP) and fluoracetamide decrease AHF,50 as do the cardiac glycosides ouabain and digoxin, which inhibit the ciliary epithelial Na+ /K+ ATPase enzyme.315 None of these have yet been shown to have any clinical relevance for the human.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


