Anatomy & Physiology of the Ear: Introduction
Mechanical events resulting from sound, gravitational forces, and rotational acceleration are detected by the cochlea and vestibular organs within the inner ear. Sound is a mechanical vibration (eg, as produced by a vibrating piano string). This vibration sets up small oscillations of air molecules that, in turn, cause adjacent molecules to oscillate as the sound propagates away from its source. Sound is called a pressure wave because when the molecules of air come closer, the pressure increases (compression); as they move further apart, the pressure decreases (rarefaction).
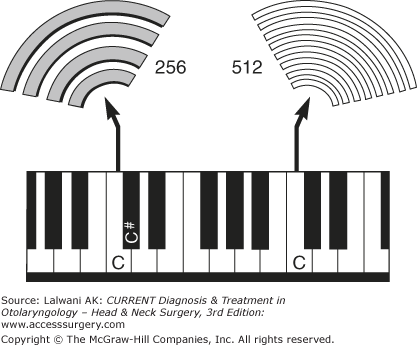
A sound is characterized by its frequency and intensity. The frequency of a sound is its pitch. Middle C on a piano has a frequency of 256 cycles per second, whereas high C (seven white keys to the right) has a frequency of 512 cycles per second (Figure 44–1). People with normal hearing can tell the difference between two sounds that differ in frequency by less than 0.5%. To appreciate how small a difference this is, one needs only to realize that middle C differs from C sharp by more than 5%. Human hearing is limited to sound waves between 20 and 20,000 Hz. Many other mammals can hear ultrasound (>20,000 Hz), and some, such as whales, approach up to 100,000 Hz.
The intensity of a sound determines its loudness and reflects how tightly packed the molecules of air become during the compression phase of a sound wave. The ear can detect sounds in which the vibration of the air at the tympanic membrane is less than the diameter of a hydrogen molecule (< 0.24 nm). The mammalian ear has the ability to discriminate a wide range of intensities—over a 100,000-fold difference in energy (120 dB).
To maximize the transfer of sound energy from the air-filled environment to the fluid-filled inner ear, land animals evolved external ears as sound collectors and middle ears as mechanical force amplifiers (Figure 44–2).
Figure 44–2.
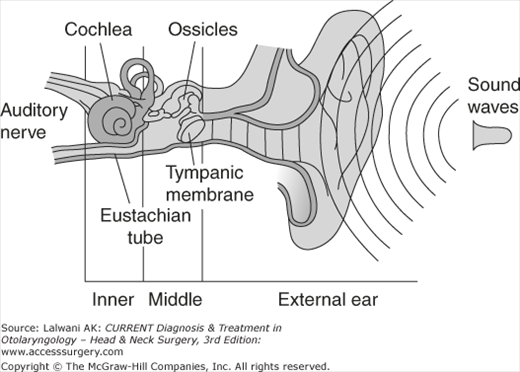
Anatomy of the ear. The external ear collects sound pressure waves and funnels them toward the tympanic membrane. The middle ear ossicles transmit the sound waves to the inner ear (cochlea). The middle ear acts to match the impedance difference between the air of the external environment to the fluid within the cochlea. This permits maximal sound transmission.
The task of the cochlea is to analyze environmental sounds and transmit the results of that analysis to the brain. The inner ear first determines how much energy is present at different frequencies that make up a specific sound. The cochlea can do this because of its tonotopic organization, whereby different frequency tones stimulate different areas of the cochlea. This mapping of frequency information is just one of the several strategies that the ear uses to code incoming information. The frequency analysis of environmental sounds begins in the external ear.
External Ear
The external ear consists of the pinna and the external auditory canal. The pinna (Figure 44–3) is a three-layered structure. The central framework consists of elastic cartilage surrounded by a layer of skin. There is minimal subcutaneous tissue between the skin and the perichondrium. Physiologically, the pinna acts to funnel sound waves into the ear canal. The intricate shape of the pinna affects the frequency response of incoming sounds differently, depending on the vertical position from which the sound originated. This information is used by the brain to localize the sound source in three-dimensional space. Overall, the shape of the external ear provides approximately 20 dB of gain to sounds in the middle frequency range (2–4 kHz).
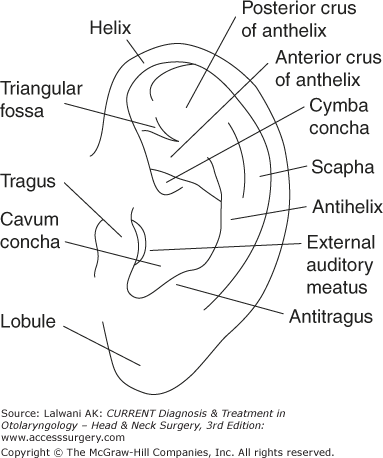
The external auditory canal consists of a lateral cartilaginous portion and a medial bony portion. Each portion of the canal takes up approximately half of its length. The tragus forms the anterior cartilaginous canal. Directly in front of it lies the parotid gland. The facial nerve exits the stylomastoid foramen 1 cm deep to the tip of the tragus (the tragal pointer). Within the anterior and inferior portions of the cartilaginous ear canal, there are small fenestrations through the cartilage called the fissures of Santorini. Infection of the ear canal (otitis externa) can spread to the parotid gland through these fissures and may lead to skull base osteomyelitis. The tympanic portion of the temporal bone forms most of the bony ear canal. Anterior to the bony canal is the temporomandibular joint. The skin of the ear canal is thicker in the cartilaginous canal and contains glands that secrete cerumen (ear wax). The skin of the bony ear canal is very thin and fixed to the periosteum. No cerumen is secreted in the bony ear canal.
The great auricular nerve (from nerve roots C2 and C3) provides sensory innervation to the skin overlying the mastoid process as well as to most of the pinna. Cranial nerves V (the trigeminal nerve), VII (the facial nerve), and X (the vagus nerve) innervate the external auditory canal.
Middle Ear
The tympanic membrane consists of three layers: outer, middle, and inner. The outer layer arises from the ectoderm, which consists of squamous epithelium. The inner layer originates from the endoderm and consists of cuboidal mucosal epithelium. The middle layer originates from the mesenchyme and is called the middle fibrous layer. The middle fibrous layer of the tympanic membrane consists of both radial and circumferential fibers. These fibers are important in maintaining the strength of the tympanic membrane as well as in aiding the proper vibration of the tympanic membrane with different frequency sounds.
The tympanic membrane has an oval shape and is approximately 8 mm wide and 10 mm high (Figure 44–4). The tympanic membrane is sloped so that the superior aspect is lateral to the inferior aspect. In addition, the tympanic membrane is tented medially by the long process of the malleus (manubrium). Around the circumference of the tympanic membrane is the fibrous annulus, which sits in the tympanic sulcus, a groove in the bone at the medial end of the external auditory canal. The annulus is incomplete superior to the anterior and the posterior malleal folds. The pars flaccida is above the anterior and posterior malleal folds while the pars tensa is inferior to the folds. The pars flaccida is also known as the Shrapnell membrane. The middle fibrous layer of the pars flaccida is weaker than that of the pars tensa. This area of the tympanic membrane can easily retract inwardly when the middle ear pressure is less than the environmental air pressure and is often the starting point of an attic cholesteatoma. Blood vessels enter the tympanic membrane through the superior external auditory canal skin (the vascular strip) as well as circumferentially from around the fibrous annulus.

The middle ear cavity (Figure 44–5) originates embryologically from the first branchial pouch. It is connected to the nasopharynx via the eustachian tube. Posterior to the middle ear cavity are the mastoid air cells, which connect with the attic through the aditus ad antrum. The middle ear cavity and mastoid air cells are lined with ciliated mucosal epithelium. Anatomically, the middle ear space can be divided into five portions based on their relationship to the tympanic annulus: the mesotympanum, hypotympanum, attic, protympanum, and retrotympanum (see Figure 44–5). The retrotympanum includes the sinus tympani and facial recess.
Figure 44–5.
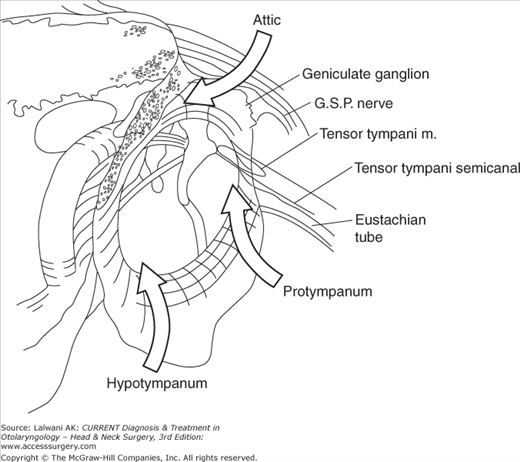
Spaces of the middle ear (a right ear). The mesotympanum is the portion of the middle ear directly behind the tympanic membrane. The attic, protympanum, and hypotympanum are superior, anterior, and inferior to the mesotympanum, respectively. The facial recess and sinus tympani are posterior to the mesotympanum (also see Figure 44–8). GSP, greater superficial petrosal.
The blood supply in the middle ear and mastoid originates from the internal and external carotid arteries. Vessels off the external carotid artery include the anterior tympanic artery and the deep auricular artery (branches of the internal maxillary artery), the superior petrosal and superior tympanic arteries (branches of the middle meningeal artery), and the stylomastoid artery (a branch of the occipital artery that runs up the stylomastoid foramen). In addition, the caroticotympanic artery, a branch of the internal carotid artery, forms a plexus over the promontory of the middle ear.
There are three ossicles (Figure 44–6): the malleus, the incus, and the stapes. The malleus has a long process, a short process, and a head. The malleus is bonded to the tympanic membrane from the tip of the long process (the umbo) to the short process. The head of the malleus articulates with the body of the incus in the attic.
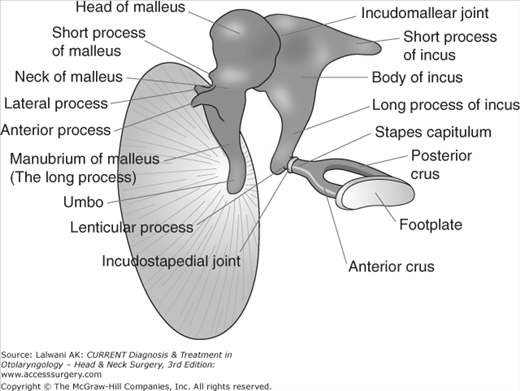
The short process of the incus is tethered to the posterior wall of the middle ear cavity for structural support and the long process is connected to the stapes capitulum. The distal portion of the long process is known as the lenticular process. The blood supply to the ossicular chain is most tentative at the lenticular process and is the first portion of the ossicular chain to be resorbed in patients with chronic otitis media, producing ossicular discontinuity.
The stapes consists of a footplate and a superstructure. The superstructure includes the anterior and posterior crus, which are attached at the capitulum. The footplate sits within the oval window.
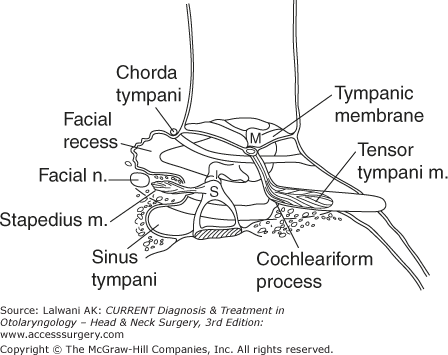
The stapedius muscle (Figures 44–7 and 44–8) originates from the pyramidal eminence. The tensor tympani muscle is anchored by the cochleariform process where it turns 90° and becomes a tendon that connects to the malleus. The ponticulus is a ridge of bone between the round window and the oval window. The subiculum is a ridge of bone just anterior to the round window. The promontory is the medial wall of the middle ear cavity. Medial to the promontory is the cochlea.
The embryologic development of the ossicles is complex. The ossicular portions found in the attic are formed from the first branchial arch. This includes the head of the malleus and the body and short process of the incus. The ossicular portions that are found within the mesotympanum originate from the second branchial arch. This includes the long process of the malleus, the long process of the incus, and the stapes superstructure. The stapes footplate originates from the otic capsule (the primordial otocyst), rather than from a branchial arch. The ossicles are full-sized cartilage models by 15 weeks of gestation, and endochondral ossification is complete by 25 weeks. The middle ear is adult sized at birth.
The facial nerve is the major nerve traversing the middle ear cavity (Figure 44–7). After entering the temporal bone via the internal auditory canal, the labyrinthine segment courses to the geniculate ganglion, immediately superior to the cochlea. The facial nerve then turns (first genu) and runs horizontally through the middle ear space (the tympanic portion of the facial nerve). The nerve lies superior to the oval window and the bone is often missing (dehiscent facial nerve) at this point. The nerve then turns again (second genu) and runs vertically (the vertical portion of the facial nerve). The nerve exits the temporal bone through the stylomastoid foramen, which is medial to the digastric muscle but lateral to the styloid process.

There are three branches of the facial nerve within the temporal bone. The greater superficial petrosal nerve (Figure 44–5) branches off at the geniculate ganglion and delivers parasympathetic nerves to the lacrimal gland and to the minor salivary glands of the nose. Another branch of the facial nerve goes to the stapedius muscle. Finally, the chorda tympani nerve branches off from the vertical portion of the facial nerve and runs underneath the tympanic membrane, medial to the malleus, before exiting the middle ear space through the petrotympanic fissure. It joins up with cranial nerve V3 and supplies both taste to the anterior two thirds of the tongue as well as parasympathetic innervation to the sublingual and submandibular glands. The cell bodies of these nerves are found in the geniculate ganglion.
Cranial nerve IX (the glossopharyngeal nerve) has a branch that runs across the tympanic promontory called the tympanic nerve or Jacobson’s nerve. It innervates the mucosa of the middle ear space and Eustachian tube as well as provides parasympathetic innervation to the parotid gland. There is also a branch of the vagus nerve within the middle ear cavity called Arnold’s nerve, which supplies innervation to the external auditory canal. Patients often cough when their ear canal is cleaned because of the referred sensation to the throat.
Understanding the area around the second genu of the facial nerve is critical for conducting safe middle ear surgery (Figure 44–8). The bony ear canal ends at the level of the annulus. The space medial to the end of the ear canal, but lateral to the facial nerve, is the facial recess. Medial to the facial nerve is another pocket of space called the sinus tympani. It is impossible to visualize the sinus tympani by looking either through the ear canal or through an opening made through the mastoid. Residual cholesteatoma is often found here because of remnants left behind (and not seen) during primary surgery.
The middle ear provides an acoustic impedance match between the environmental air and the fluid-filled inner ear. The middle ear amplifies the airborne sound vibration in two ways. First, the large surface area of the tympanic membrane, compared with the small surface area of the stapes (14:1), imparts an increase in vibrational amplitude. Second, the lever arm effect of the malleus and incus imparts a further increase in vibrational amplitude (1.3:1.0). Thus, the total middle ear gain is between 20 and 35 dB. In addition, the mass and stiffness of the ossicular chain affect its frequency response. Overall, the middle ear acts as a band-pass filter, with a maximum energy transfer over the range of 1–10 kHz.
Changing the mass and stiffness of the middle ear modulates its frequency response, which can be observed clinically. For example, the stapedius and tensor tympani muscles contract through a neural reflex arc mediated by loud sounds (>80 dB). They act to stiffen the ossicular chain and protect the inner ear from noise damage, particularly at low frequencies. In contrast, cholesteatoma formation in the middle ear can contact the ossicular chain, increasing the total mass, causing a predominantly high-frequency conductive hearing loss.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


