Anatomy of the Uveal Tract
Dimple Modi
Deepak P. Edward
The uvea is derived from the Latin root, uva, meaning grape. The uveal tract consists of a pigmented, highly vascular loose fibrous tissue that can be divided into three anatomical regions: anterior iris, central ciliary body, and the posterior choroid.1 The uveal tract is firmly adherent to the scleral spur anteriorly, the optic disc posteriorly, and at the exit points of the four vortex veins in between. The uvea provides nutrients to the retina and lens as well as assists in refractive function of the eye.
Embryologically, most of the uveal tissue develops from neural crest cells.2 The uveal vasculature develops as early as the optic vesicle stage with the formation of a large plexus of primitive vessels that originate from the neural tube vascular system and extend around the outer layer of the optic cup.3 Its blood supply comes from the ophthalmic artery; vascular and nerve supply are present both anteriorly via the anterior ciliary vessels and nerves (Fig. 1) and posteriorly via the posterior ciliary nerves and vessels (Fig. 2). A complex arrangement of smooth muscle is part of the anterior and central portions of the uvea, controlling pupillary size and accommodation, respectively.
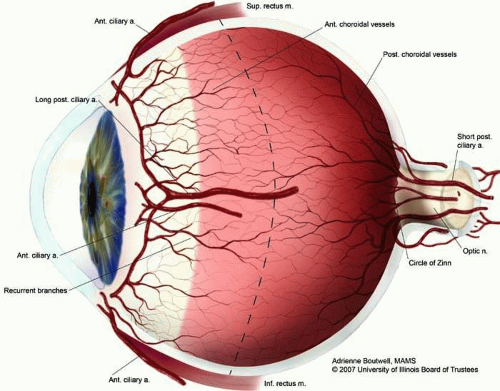 Figure 1. Vascular supply of the uveal tract. |
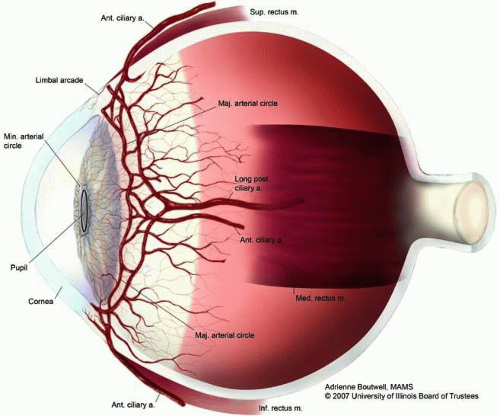 Figure 2. Vascular supply at the major arterial circle of the iris. |
IRIS
General Anatomy 1
The iris is the most anterior portion of the uveal tract. It divides the anterior segment of the eye into the anterior and posterior chambers and is bathed by aqueous on both sides. The iris is a continuous structure but histologically can be divided into multiple layers as discussed below.
Table 1: Iris Embryology | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
The precursors of the iris—the mesoderm and neuroectoderm—appear by the sixth week of gestation. At eight to ten weeks gestation, the inner layer of the anterior portion of the optic cup, which is analogous to the posterior epithelial layer of the iris, is nonpigmented; pigmentation can be observed by ten weeks. The sphincter muscle arises from the anterior layer of the neuroepithelium and is present at 11 to 12 weeks gestation. The dilator muscle develops after 23 weeks gestation. The iris vascular system develops after the eighth week of gestation. Specifically, the stroma is derived from neural crest cells. The iris musculature is derived from the outer lamina of the optic cup and the pigment epithelium from the inner optic cup (neuroepithelium). Therefore, the posterior iris, where Schwalbe’s furrows are located, is formed from the anterior portion of the primitive optic cup.
Macroscopic Anatomy4
The iris measures approximately 12 mm in diameter and 38 mm in circumference, with its free 3-mm border surrounding the pupil. The pupil pierces the iris diaphragm slightly below and nasal to its center. The pupillary zone is located between the collarette and the pupillary ruff. The mobility of the iris allows the pupil to change size. When the pupil dilates, the iris has numerous folds; during miosis, the anterior surface of the iris is smooth. The thinnest portion of the iris is at its root (0.5 mm); therefore, blunt trauma leading to iridodialysis most commonly affects the iris root while the thick collarette (0.6 mm) remains intact. This thickest portion, the collarette, is a circular ridge approximately 1.5 mm from the pupillary margin.
The anterior surface of the iris is richly textured and thin. Radial, collagenous columns or trabeculae are evident in lightly pigmented irises, although can be masked in darker pigmented individuals due to increased iris pigmentation. Thicker, radially oriented, branching trabeculae encircle depressions or openings in the surface called crypts. Crypts are located on both sides of the collarette. The collarettes are a series of trabeculae that lie approximately 1.6 mm from the pupil margin. They divide the iris surface into an outer ciliary zone and inner pupillary zone and allow for quick entry and exit of the aqueous into spaces in the iris stroma. Defects in the superficial layer of the iris form crypts, or large pit-like depressions, termed Fuchs’ crypts. The posterior surface of the iris is a more uniform, smooth, dark-brown layer.
Iris Color
Three main factors in the structure of the iris may influence its color: (1) the pigment epithelium; (2) the density of the iris stroma; and (3) the pigment content of the iris stroma. The iris pigment epithelium pigmentation does not vary between different iris colors, and clump cells of histiocytic origin are relatively few in number. Various shades of iris color have been attributed to the variability in number and distribution of melanin granules within superficial stromal melanocytes.5 There are two types of melanocytes—eumelanin which appears brown or black and pheomelanin which results in yellow or red pigment. The iris stroma mainly consists of eumelanin aggregated in the anterior leaf of the iris. A study by Albert et al indicated that the mean melanocyte numbers found between Caucasians and African American irides were similar but that Asian irides had fewer total melanocytes and total number of cells than the former two groups.6
Microscopic Anatomy7
There are four contiguous layers that compose the iris: (1) the anterior border layer; (2) the stroma and sphincter muscle; (3) the anterior iris pigment epithelium and dilator muscles; and (4) the posterior iris pigment epithelium (Fig. 3).
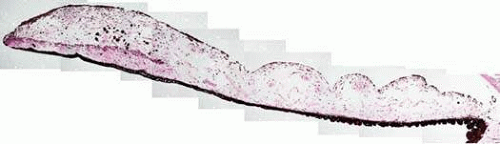 Figure 3. Iris at low magnification showing the anterior border layer, stroma, iris musculature and layers of iris pigment epithelium (hematoxylin and eosin; original magnification 3×). |
Anterior Border Layer
The anterior layer of the iris, the anterior border layer, consists of a condensation of connective tissue, fibroblasts, and melanocytes derived from the anterior stroma. The border layer is absent at crypts and thinned at contraction furrows. It is thickest in the pupillary zone and at the periphery of the ciliary zone.
The fibroblasts which are the predominant cells in this layer line the many ridges and crypts, stretching from the iris root to the pupil. Aqueous humor flows freely through these crypts and crevices. Numerous small folds extend radially for 1 mm from the edge of the pupil, forming Schwalbe’s contraction folds in the posterior iris surface. These Schwalbe’s structural folds involve both epithelial and vascular layers; they begin 1.5 mm from the pupil margin and pass into the valleys of the ciliary processes. Circular furrows cross these structural folds at regular intervals and are due to variation in the thickness of the pigment epithelium.
Pigmented melanocytes lie deeper to the fibroblasts. The distribution of melanocytes within this layer may vary throughout the iris. Gap junctions, intermediate junctions, and tight junctions are all found to connect cells of similar phenotype within the anterior border layer. The exception is gap junctions, which have been shown to exist between fibroblasts and melanocytes. This layer also contains capillaries and venules as well as nerve fibers that terminate in the adjacent stroma.
Iris Stroma and Sphincter Muscle
The stroma is composed of a loose network of collagenous tissue; amidst this tissue are pigmented and nonpigmented cells, collagen fibrils, and extensive ground substance (Fig. 4). The pigmented cells include melanocytes and clump cells, whereas the nonpigmented cells consist of fibroblasts, lymphocytes, macrophages, and mast cells. Wide spaces within the stroma prevent formation of a meshwork between the branching processes of melanocytes and fibroblasts. They also allow for diffusion of aqueous as well as facilitate pupil movement. Clump cells are large round darkly pigmented cells found in the stroma of mammalian irises. These cells are also known as clump cells of Koganei and are characteristically found in the pupillary region near the sphincter muscle and in the anterior ciliary body near the iris root. The fibril network anchors vessels in place within the stroma and protects them from bending during excessive iris movement with miosis and mydriasis. The iris stroma is continuous with the stroma of the ciliary body.
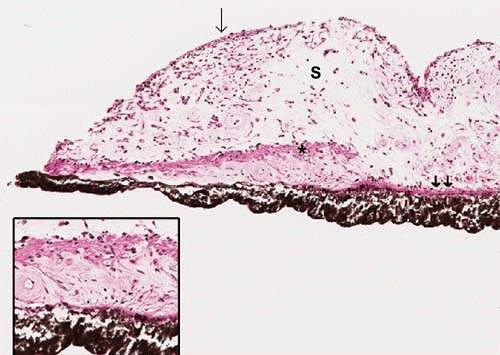 Figure 4. Iris at the pupillary zone. Note the anterior iris border layer (arrow), iris stroma (S); sphincter muscle (*); dilator muscle (double arrow). Inset: Note the double layered iris pigment epithelium and the dilator muscle that appears continuous with the anterior layer of the iris pigments epithelium. Also note the spindle shaped melanocytes in the stroma (original magnification 5×; Inset 18×) |
The sphincter muscle lies within the deep stroma and is composed of smooth muscle cells joined by tight junctions. The sphincter muscle is anchored firmly to adjacent stroma. Contraction of the sphincter muscle causes the pupil to constrict in miosis. The sphincter muscle retains its function even if severed radially. Therefore, if the sphincter muscle is cut during a sector iridectomy (where a wedge shape of iris including the tissue at the pupillary border is removed), the ability of the muscle to contract is not lost. The sphincter muscle is further discussed in the “Musculature” section below.
Anterior Iris Pigment Epithelium and Dilator Muscle
There are two layers of iris pigment epithelium posterior to the stroma. The anterior iris epithelium closer to the stroma is composed of myoepithelial cells (Fig. 4, inset). These cells are unique because the apical portion of the cell is pigmented cuboidal epithelium joined by tight junctions and desmosomes, whereas the basal portion is composed of elongated contractile smooth muscle processes. These muscle fibers extend into the stroma, forming the dilator muscle. Small projections of this muscle insert into the sphincter and contraction of these radially arranged fibers enlarges the pupil causing mydriasis. The dilator muscle is further discussed in the “Musculature” section below.
Posterior Iris Pigment Epithelium
The deeper epithelial layer posterior to the stroma is the posterior iris epithelium. Here, a single layer of heavily pigmented columnar cells are joined by tight junctions and desmosomes. In the periphery, the posterior iris epithelium begins to lose its pigment as it continues into the ciliary body as nonpigmented epithelium. The apical microvilli extend from surfaces of both the posterior and anterior epithelia and interdigitate, with scattered 20-nm spaces between them; desmosomes join these two apical surfaces. The anterior and posterior epithelial layers are arranged apex to apex in two layers, absorbing incident light that does not enter through the pupil.
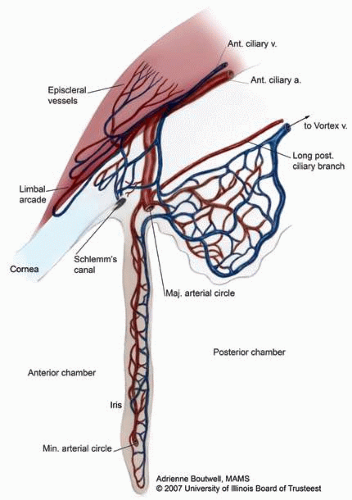 Figure 5. Vascular supply of the iris. |
Blood vessels form the bulk of the iris stroma. The vascular supply to the iris originates in the anterior and long posterior ciliary branches of the ophthalmic artery (Fig. 6). These branches merge at the ciliary body to form the major arterial circle before entering radially into the iris. The arterial circle is formed by the junction of the long posterior ciliary arteries and the anterior ciliary arteries that extend from the insertion of the extraocular rectus muscles. Because the anterior ciliary artery branches extend from all rectus muscles except the lateral, care should be taken during muscle surgery to avoid anterior segment ischemia.
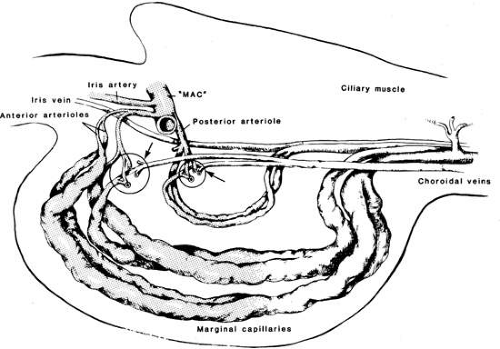 Figure 6. The vasculature at the iris root and ciliary body. Note the vascular communications between the iris vasculature and the choroid (From Morrison JC, Van Buskirk EM: Ciliary process microvasculature of the primate eye. Am J Ophthalmol 97:372–383, 1984, with permission). |
An incomplete circular vessel, the minor circle of the iris, is located in the iris stroma in the region of the collarette and is a remnant of embryologic development. The minor vascular circle of the iris is formed by anastomoses between the arterial and venous arcades. Arteries in the iris are often larger than veins and the veins have no valves. The vessels lack an internal elastic lamina and the iris capillaries are lined by nonfenestrated endothelial cells to form part of the blood-aqueous barrier. A layer of loose connective tissue surrounds the iris capillaries.
Nerves7
The oculomotor nerve (cranial nerve III) is involved in sympathetic and parasympathetic innervation of the iris. The parasympathetic, sympathetic, and sensory nerve fibers all run through the ciliary ganglion, but only parasympathetic fibers form synapses in the ganglion.
The presynaptic parasympathetic fibers originate in the Edinger-Westphal nucleus, the parasympathetic motor nucleus associated with the oculomotor nucleus in the brainstem. Axons from the Edinger-Westphal nucleus and the oculomotor nucleus run together in the brainstem and exit together as the oculomotor nerve. Postsynaptic parasympathetic fibers leave the ciliary ganglion in multiple (six to ten) short ciliary nerves. These nerves enter the posterior aspect of the eyeball and travel forward to innervate the sphincter pupillae and ciliary muscles.
The sympathetic root of the ciliary ganglion contains postganglionic sympathetic fibers whose cell bodies are located in the superior cervical ganglion. Their axons ascend with the internal carotid artery as a plexus of nerves, the carotid plexus. Sympathetic fibers innervating the eye separate from the carotid plexus within the cavernous sinus. They run forward through the superior orbital fissure and merge with the long ciliary nerves (branches of the nasociliary nerve) and the short ciliary nerves (from the ciliary ganglion). Sympathetic fibers in the short ciliary nerves pass through the ciliary ganglion without forming synapses. Sympathetic fibers from the superior cervical ganglion innervate the dilator pupillae.
Sensory fibers from the iris run posteriorly through the short ciliary nerves and pass through the ciliary ganglion without forming synapses. They leave the ciliary ganglion in the sensory root, which joins the nasociliary nerve. The nasociliary nerve is a branch of the ophthalmic nerve, one of the three branches (V1) of the trigeminal nerve. The trigeminal nerve is the main sensory nerve of the face.
The smooth muscle and pigment epithelial structures are located in the posterior portion of the iris. The pupillary sphincter is formed by smooth muscle tightly arranged in a circle anterior to the pigment epithelium of the iris. It is a flat muscular strap 0.75 mm wide and 0.1 to 0.17 mm thick. Each fusiform smooth muscle cell within the sphincter is enveloped in a basal lamina. These cells are grouped in bundles of five to eight cells with gap junctions between them to facilitate the spread of depolarizing current within a muscle group.
The muscular processes of the dilator muscle measure up to 60-μm long and 7-μm wide. These dilator muscle processes extend from their cell bodies in the anterior pigment epithelial layer. Close to the pupil margin, the dilator muscle fuses with the deep surface of the sphincter muscle. When the muscle contracts, it leads to mydriasis. The dilator muscle is innervated by the sympathetic nervous system via the long ciliary nerves.



