Fig. 1.1
Membranous labyrinth of vertebrates. Membranous labyrinths of a rat (a), a bird (b), and a fish (c) were shown. (Modified from “The Vertebrate body” by Romer et al. [1])
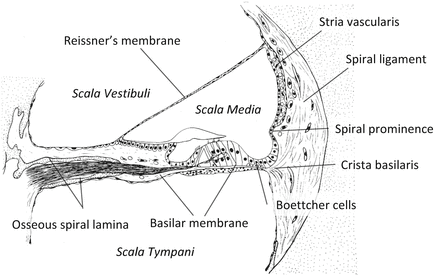
The axis of the cochlea is known as the modiolus, in which cochlear nerve runs (Fig. 1.2). The cross section of the membranous labyrinth in the cochlea forms triangular shape bordered by the basilar membrane, the stria vascularis, and the Reissner’s membrane (Fig. 1.3). The endolymphatic space inside the cochlea is called scala media or cochlear duct. The perilymphatic space facing the Reissner’s membrane is called scala vestibuli, and the space facing the basilar membrane is called scala tympani. The scala tympani and scala vestibuli are connected through a small hole of the basilar membrane at the apex of the cochlea, called a helicotrema. Therefore, perilymphatic space is divided into two spaces by the cochlear duct. The basal portion of the scala tympani ends with a blind sac. The basal end of the scala vestibuli is connected to the saccule via ductus reuniens. The basilar membrane is a resonant structure spreading between osseous spiral lamina and the crista basilaris. The basilar membrane in the basal turn is narrower and stiffer, leading to higher natural resonance frequency, than that in the apical turn. The organ of Corti is positioned on the scala media side of the basilar membrane.
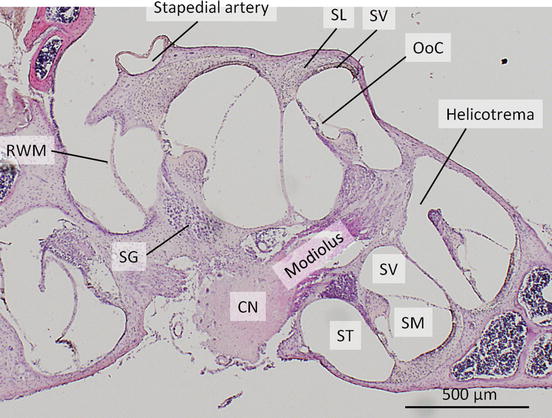
Fig. 1.2
Cochlea. Mid-modiolar section of an adult mouse cochlea, stained with hematoxylin and eosin. The cochlear nerve (CN) runs in the modiolus. Cell bodies of CN exist as spiral ganglions (SG) in Rosenthal’s canal. The cochlear duct is seen as scala media (SM). Scala tympani (ST) and scala vestibuli (SV) are connected in the apical turn of the cochlea as helicotrema. Stapedial artery runs on the surface of the basal turn of the cochlea. Round window membrane (RWM), the organ of Corti (OoC), stria vascularis (SV), the spiral ligament (SL)
The organ of Corti (Figs. 1.2, 1.3, and 1.4) is composed of hair cells and several types of supporting cells. When the sound vibration displaces the basilar membrane, the shearing force between the basilar and the tectorial membranes deflects the hair bundle at the apical surface of the inner hair cells and then is conveyed as the release of the neurotransmitter. Approximately 3,500 inner hair cells exist in one human cochlea [5]. The inner hair cells are innervated by type I afferent neurons with myelinated axons. The inner phalangeal cells are the direct supporting cells for inner hair cells. Border cells and inner sulcus cells are located next to the inner phalangeal cells. The outer hair cells appear as three rows, and 12,000 outer hair cells exist in one human cochlea [5]. These cells change their length through the function of the motor protein, prestin, in response to the sound stimuli [6, 7]. The outer hair cells are innervated by type II afferent neurons with unmyelinated axons. Deiters’ cells support the outer hair cells. Deiters’ cells have two parts: The cell body directly attaches to the basilar membrane and supports the basal side of the outer hair cell. The phalangeal process extends from the cell body to the apical surface of the sensory epithelium between outer hair cells and forms a part of reticular lamina. Inner and outer pillar cells exist between inner hair cells and the first row of outer hair cells and form a fluid-filled space called the tunnel of Corti. The space between outer pillar cells and the first row of outer hair cell/Deiters’ cell is called Nuel’s space. Hensen’s cells are located next to the third row of the Deiters’ cells and form several layers. Claudius’ cells exist between Deiters’ cells and the spiral prominence, forming outer sulcus. Boettcher cells are located beneath the Claudius’ cells in the lower turns of the cochlea (Fig. 1.3). Tympanic border cells are cells lining the scala tympani side of the basilar membrane.
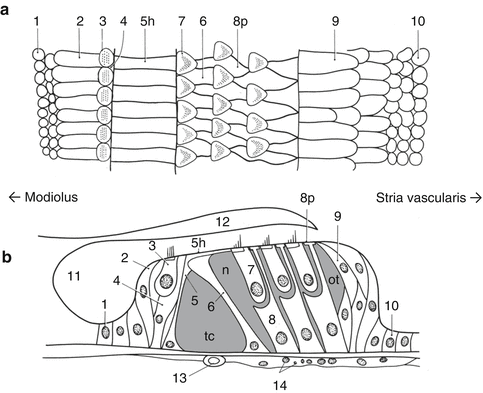
Fig. 1.4
The organ of Corti, surface view (a) and cross sectional view (b). The tunnel of Corti (tc), Nuel’s space (n), and the outer tunnel (ot) are filled with cortilymph. 1 Inner sulcus cell, 2 border cell, 3 inner hair cell, 4 inner phalangeal cell, 5 inner pillar cell, 5h the head of inner pillar cell, 6 outer pillar cell, 7 outer hair cell, 8 Deiters’ cell, 8p phalanx of Deiters’ cell, 9 Hensen’s cell, 10 Claudius’ cell, 11 inner spiral sulcus, 12 tectorial membrane, 13 vessel of the basilar membrane, 14 tympanic border cell. (Modified from “Atlas of Otology” by Nomura et al. [5])
The loose connective tissue on the outer wall of the cochlear duct is the spiral ligament (Fig. 1.3). The stria vascularis is a vascular-rich structure on the spiral ligament spreading from the attachment of Reissner’s membrane to the spiral prominence. The stria vascularis functions to maintain the high potassium concentration and positive potential of the endolymph (endocochlear potential (EP)) via Na/K-ATPase.
The basilar papilla is a sensory epithelium in the cochlear duct of birds, amphibians, and lizards, which functions like the organ of Corti (Fig. 1.5) [3]. Hair cells and supporting cells are compactly arranged on the basilar membrane. The tall hair cells exist near the superior margin of the basilar membrane, while the short hair cells exist near the inferior margin.
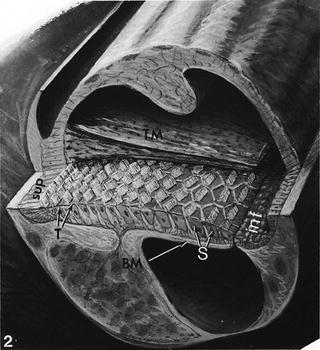
Fig. 1.5
The basilar papilla of a chicken. T tall hair cell, S short hair cell, inf inferior margin of the cochlea, sup superior margin of the cochlea, BM basilar membrane, TM tectorial membrane. (From Tilney and Saunders [3])
1.1.2 The Vestibule
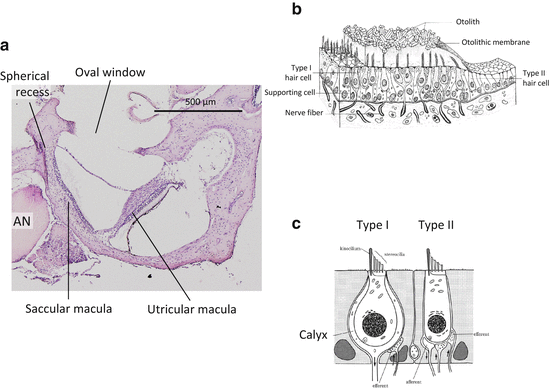
The vestibule (the utricle and saccule) is positioned superior to the cochlea and functions to sense linear acceleration (Fig. 1.6). In the vestibule, the membranous labyrinth forms two sacs which are connected each other. The sac near the semicircular canals is called the utricle, while the other located near the cochlea is called the saccule. The saccule is connected to the cochlea by a small channel called the ductus reuniens. The sensory epithelium of the vestibule is located in the macula of the utricle and saccule. The utricular macula is located horizontally with a free margin. It resembles a visor of a cap separating the utricle and perilymphatic space of the vestibule. The saccular macula is attached to the medial wall of the saccule. The bony labyrinth forms a shallow bowl called spherical recess at the attachment site of saccular macula. Each macula is overlaid by the otolithic membrane, which is a rigid layer composed of gelatinous extracellular matrix and otolith. The space between the otolithic membrane and the apical surface of the sensory epithelium is secured by the columnar filament layer [8]. Each macula is divided into two regions by the striola. In the utricle, hair cells are arranged in such a manner that the tall cilia face the striola. In the saccule, the tall cilia face the peripheral side instead of the striola. Hair cells in the maculae are classified into type I and II hair cells [5, 9]. Type I hair cells have a bulbous shape and its basal part is surrounded by large socket-like afferent nerve endings called the calyx. Type II hair cells have a cylindrical shape and display simpler button-like afferent nerve contacts. Both type I and II hair cells receive efferent nerve contacts. Type I hair cells exist near the striola, and type II in the peripheral region. Type I and II hair cells are morphologically distinguishable, but the functional difference is not fully understood. Dark cells are responsible for the maintenance of endolymph through ion and water transport [10]. Dark cells exist around the edge of the utricular macula, but not the saccular macula [11].