1 An Introduction to Ocular Anatomy and Histology
A paired, hollow, spherical organ, the human eye is slightly less than an inch in diameter (Fig. 1-1A). Internally, it is divided into anterior and posterior chambers and a much larger vitreous cavity, which comprises most of its posterior segment. The anterior chamber is bounded anteriorly by the cornea and posteriorly by the anterior surfaces of the iris and lens (Fig. 1-1B). The posterior chamber is bounded anteriorly by the iris pigment epithelium (IPE), laterally by the ciliary processes that form the pars plicata of the ciliary body, and posteriorly by the “face” or anterior surface of the vitreous humor. Both anterior and posterior chambers are filled with watery aqueous humor, which is secreted by the ciliary epithelium on the ciliary processes.
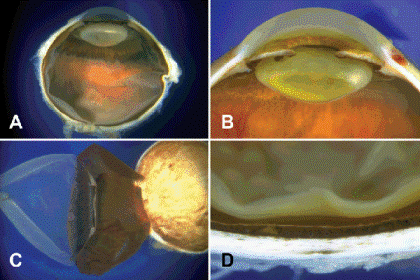
Fig. 1-1. A. Slightly less than an inch in diameter, the eye is composed of an anterior chamber and a much larger posterior vitreous cavity. B. The anterior chamber is bounded anteriorly by the cornea and posteriorly by the iris and lens. The lens is situated in the posterior chamber behind the pupil. C. The eye is composed of three concentric coats that are easily separated during dissection. D. Posteriorly, the three coats are evident as the sclera, pigmented choroid, and the retina.
The vitreous humor fills most of the posterior segment. The most delicate connective tissue in the body, the vitreous humor is a transparent gel composed of hyaluronic acid and a framework of delicate fibrils of type II collagen. Centrally, the anterior face of the vitreous abuts the posterior surface of the lens, which is located directly behind the iris in the posterior chamber, and is supported by a suspensory ligament of zonular fibers. The concavity in the anterior face of the vitreous that holds the lens is called the patellar fossa, and the retrolental space of Berger is interposed between the lens and the vitreous. Posteriorly, the vitreous lines the inner surface of the retina. Anteriorly, it is firmly adherent to the vitreous base, which straddles the ora serrata, the junction between the peripheral retina and the pars plana of the ciliary body. The vitreous also has relatively firm attachments to the margin of the optic disc and major retinal vessels. A curved channel called Cloquet canal runs through the center of the vitreous from the lens to the optic nerve. Overlying the optic nerve head, the posterior part of Cloquet canal widens to form a space resembling an inverted funnel called the area Martegiani.
The eye wall is composed of three concentric coats that vary markedly in their cellularity and composition (Fig. 1-1C,D). The three layers are readily separated during gross dissection. The outer coat includes the white sclera that merges with the transparent cornea at the corneoscleral limbus. Paucicellular and composed largely of dense collagenous connective tissue, the outer layer constitutes the eye’s tough fibrous exoskeleton that confines and protects its contents of fluid and delicate neural tissues. The collagenous lamellae of the sclera are larger and more irregular than that of the cornea.
The eye’s innermost coat is composed of neuroectodermal tissues derived from the optic cup. Highly cellular and largely devoid of connective tissue, these structures include the neurosensory retina, the retinal pigment epithelium (RPE), the ciliary epithelium, and the IPE. Embryologically, this inner layer of neuroectodermal tissues is derived from an outpouching of the forebrain called the optic vesicle. The neuroectoderm is a polarized layer of epithelial cells with characteristic apical and basal features. Microvilli and cilia typically are found on the apical surface of the cells, which are joined near their apices by intercellular connections called terminal bars. The bases of the cells rest on a basement membrane or basal lamina. The basal lamina serves as the attachment for extracellular matrix material.
The optic vesicle is lined externally by a basement membrane, and the apical surface of its neuroectodermal cells projects into its lumen. Later, as the optic vesicle invaginates to form the optic cup on day 27 of gestation, the apical surfaces of the inner and outer layer of cells become approximated. This “apex to apex” orientation continues throughout life. The inner layer of the optic cup goes on to form the posterior layer of IPE, the inner nonpigmented layer of ciliary epithelium, and the neurosensory retina. Its outer layer forms the anterior half of the IPE, which includes the dilator muscle, the outer pigmented layer of ciliary epithelium, and the RPE.
The two layers of cells that compose the iris and ciliary epithelia retain “apex to apex” orientation found in the optic cup with basal laminae on their external surfaces. Both layers are firmly fused together. Both layers are pigmented in the IPE. The ciliary epithelium is a half-pigmented bilayer; its outer layer is pigmented and its inner layer is nonpigmented (see Fig. 1-5D). During microscopy, this “two-toned” bilayer of cells readily serves to identify ciliary body tissue. At the ora serrata, the outer layer of pigmented ciliary epithelium continues posteriorly as the RPE, and the inner layer of nonpigmented ciliary epithelium abruptly thickens to form the neurosensory retina, with its complex lamellar architecture. Posterior to the ora serrata, the layers derived from the inner and outer layers of the optic cup are not fused. A potential space called the subretinal space is present between the retina and the RPE. The apical surfaces of both tissues project into the subretinal space and interdigitate. Retinal detachment or separation is marked by the accumulation of fluid in this potential space.
In routine microscopic sections, the IPE appears to be a single layer, since its two layers of cells are obscured by heavy pigmentation (Fig. 1-2D). Both layers are apparent in bleached sections, however. The iris dilator muscle appears as an eosinophilic band on the anterior surface of the IPE. The dilator is not a separate structure. Rather, it is composed of cellular processes arising from the basal surface of the anterior layer of cells, which are nonpigmented and have smooth muscle differentiation. The sphincter muscle of the iris, which encircles and constricts the pupil, is also derived from the IPE but separates from it during development, forming a distinct structure. Continuity of the sphincter muscle with the IPE can be observed microscopically in fetal eyes. The sphincter is about 1 mm in width and is separated from the IPE by a layer of connective tissue that becomes more prominent with increasing age.
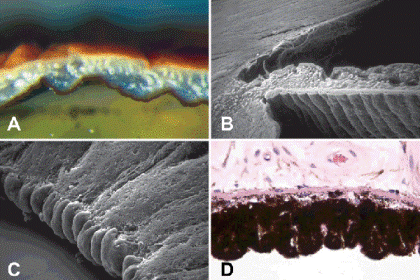
Fig. 1-2. The iris. A. The pigment in this brown iris is concentrated in the melanocytes of the anterior border layer. The posterior IPE is maximally pigmented. Cuffs of collagen highlight iris stromal vessels. B. SEM shows circumferential ridges on posterior surface of the neuroectodermal IPE. Iris stroma is derived from the neural crest. C. Termination of the IPE at pupillary margin forms beaded pigment ruff. D. The two layers comprising the IPE are fused and maximally pigmented. The dilator muscle is evident as an eosinophilic band on its stromal surface. It is composed of cellular processes with smooth muscle differentiation. (B. SEM ×40, C. SEM ×320, D. H&E ×250)
Clinically, the IPE is visible at the edge of the pupil as the pigment ruff or frill, which corresponds to the margin of the optic cup (Fig. 1-2C). This is the only neuroectodermal tissue of the eye that can be seen with the slit lamp biomicroscope without supplementary lenses. The granules of neuroepithelial melanin in the iris pigment epithelial cells are large and spherical in shape in contrast to RPE granules, which typically are ellipsoidal. Granules of neuroectodermal melanin are always larger than the dustlike melanosomes found in uveal stromal cells. The IPE is maximally pigmented in all eyes.
A peripheral colony of brainlike tissue, the neurosensory retina is a complex, highly cellular tissue composed of cells that are arranged in regular layers (Fig. 1-3A). Ten retinal layers including the RPE are identified. It is relatively easy to remember the microscopic anatomy of the retina if one recalls that the retina is a three-neuron system composed of three layers of cells: photoreceptors and first- and second-order neurons. Regarding orientation, the term inner refers to the layers of cells that are located closest to the vitreous cavity or the center of the eye. Outer denotes layers that are located nearer to the sclera. The nuclei of retinal cells are arranged in distinct bands called nuclear layers. The nuclei of the photoreceptors and first-order neurons (bipolar cells) form the outer nuclear layer (ONL) and inner nuclear layer (INL), respectively. The retinal ganglion cells (the second-order neurons) comprise the innermost layer of nuclei in the retina. The ganglion cell layer (GCL) varies markedly in thickness. Surrounding the fovea, the GCL is five or more cells thick. Multilamination of the GCL is an excellent histologic marker for the macula or perifoveal part of the retina. The GCL is thin in the peripheral retina. Peripheral to the vascular arcades formed by the superior and inferior temporal retinal arterioles, the ganglion cells form a discontinuous monolayer.
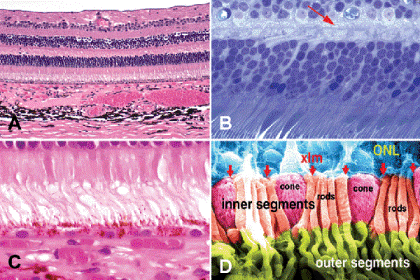
Fig. 1-3. The retina. A. The retina is composed of orderly layers of cell nuclei (nuclear layers) and plexiform layers composed of axons and dendrites. The photoreceptors rest on the RPE on the inner surface of the choroid. B. The MLM (arrow) is a linear band of synapses connecting photoreceptor cell axons and bipolar cell dendrites in the anterior part of the OPL. C. The inner and outer segments of the photoreceptors project through the fenestrated XLM into the subretinal space. The tips of the outer segments interdigitate with pigmented processes on the apical surface of the RPE. D. False colorized SEM highlights inner and outer segments of rods and cones. Cones are distinguished by the conical configuration of their inner segments. Photoreceptor nuclei comprise ONL. (A. H&E ×50, B. Toluidine blue ×250, H&E ×250, SEM ×2,500)
The plexiform layers of the retina are composed of axons and dendrites. The outer plexiform layer (OPL) is interposed between the ONL and INL, and the inner plexiform layer separates the INL from the GCL. The inner plexiform layer is truly plexiform: its whole breadth is comprised of an intricately interweaving tangle of ramifying neuronal processes. In contrast, only the narrow inner band of the OPL is truly plexiform. Its wider outer part contains an orderly parallel array of photoreceptor axons called Henle fibers. Although Henle fibers occur throughout the retina, the eponym generally is applied to the radially oriented photoreceptor axons surrounding the fovea. A line of synaptic connections comprising cone pedicles, rod spherules, and the dendrites of bipolar cells delimits the outer margin of the truly plexiform part of the OPL (Fig. 1-3B). This linear row of synapses is called the middle limiting membrane (MLM). Like the retina’s external limiting membrane (XLM), the MLM is not a basement membrane (the only true basement membrane of the retina is the internal limiting membrane or ILM). The MLM delimits the vascularized inner part of the retina; capillaries from the central retinal artery penetrate no deeper than the MLM. External to the MLM, the retina is avascular and depends on the choroidal circulation for oxygen and nutrients. The OPL is a watershed zone between the dual vascular supplies of the retina. This factor contributes to the localization of edema fluid and hard exudates in OPL.
The INL contains the nuclei of bipolar, amacrine, horizontal, and Mueller cells. Bipolar cells predominate. The horizontal cell nuclei are found in the outer part, and the amacrine cell nuclei in the inner part of the INL. The nuclei of the Mueller cells can lie at any level of the INL. Accessory glial cells including fibrous and protoplasmic astrocytes and oligodendrocyte-like cells are confined to the nerve fiber, ganglion cell, and inner plexiform layers of the retina. Hence, gliosis does not occur after central retinal artery occlusion because these cells are killed.
As we have seen, the orientation of cells in the optic vesicle and cup persists in the adult eye. The internal limiting membrane (ILM) of the retina corresponds to the basal lamina on the inner layer of the optic cup. A true basement membrane, the ILM is synthesized by the basal foot processes of the giant glial cells of Mueller, which span the entire thickness of the retina. The ILM serves as an attachment point for the fine fibrils of type II collagen that comprise the fibrillar component of the vitreous humor. Intricately interweaving processes of the Mueller cells totally fill the interstices between the retinal neurons and serve to segregate their receptive surfaces. There is little or no extracellular space in the retina.
The Mueller cells are polarized. Electron microscopy discloses microvilli on their apical surfaces, which project into the subretinal space between the photoreceptors. A belt desmosome of intercellular junctions called the XLM is found in the outer or apical part of the neurosensory retina. Not a true basement membrane, the XLM is composed of permeable adherent junctions (zonulae adherentes) that join the apices of the Mueller cells and the rod and cone cells. The XLM is permeable and does not form a barrier to fluid or macromolecules. The XLM has been called the fenestrated membrane of Verhoeff. The inner and outer segments of the rods and cones project through this fenestrated membrane into the subretinal space where they are surrounded by extracellular matrix material rich in hyaluronidase-resistant acid mucopolysaccharide (Fig. 1-3C). Cone cell nuclei are located next to the XLM in the ONL.
Rods and cones are distinguished light microscopically by the shape of their inner segments (Fig. 1-3D). The inner segments of rods are slender rodlike cylinders, while extrafoveal cones are conical in shape. Highly specialized cones that resemble rods are densely packed in the cone-rich fovea. The photoreceptor inner segments are packed with mitochondria that provide energy for the photochemistry of vision, which occurs in the photoreceptor outer segments. Photoreceptor inner and outer segments are joined by a connecting cilium that has the characteristic 9 + 0 pattern of microtubular doublets found in central nervous system cilia. Photoreceptors probably are derived from modified cilia, which are found on the apical surface of cells.
The outer segments of the photoreceptors contain stacks of thin discs composed of cellular membranes. The arrangement of the membranous discs has been likened to a stack of coins or potato chips packaged in cylindrical containers. Visual pigments such as rhodopsin are incorporated as transmembrane proteins in the disc membranes. Separate, free-floating discs occur in rods. The discs in cone outer segments are not separate structures; they comprise a continuous, sinuously folded structure formed by invaginations of the outer cell membrane.
Photoreceptor outer segments project from the apical surface of the retina into the subretinal space where they are enveloped by the microvillous processes on the apical surface of the RPE, the polarized monolayer of pigmented cells derived from the outer layer of the optic cup. The RPE cells are firmly joined near their apices by a girdle of intercellular connections called zonulae occludentes. These tight junctions form a barrier to the passage of molecules and constitute the outer part of the blood–retinal barrier. Large ellipsoidal granules of melanin pigment measuring approximately 1 µm in diameter are found in the apical cytoplasm of the RPE cells. Granules of lipofuscin pigment fill the basal cytoplasm. This “wear and tear” pigment is derived from the incomplete digestion of photoreceptor outer segments by the RPE’s phagolysosomal system and gradually accumulates with age. The lipofuscin is relatively inapparent in routine Hematoxylin and eosin (H&E) sections, but the pigment is autofluorescent and is vividly disclosed by UV fluorescent microscopy (Fig. 1-4D). The nuclei of the RPE cells are located in the basal cytoplasm near Bruch membrane.
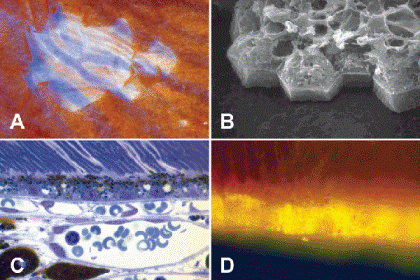
Fig. 1-4. The retinal pigment epithelium (RPE). A. The RPE is largely responsible for the color of the fundus. Rip in pigmented layer discloses large vessels in underlying choroid separated by uveal pigment. B. SEM shows hexagonal monolayer of RPE cells covering the inner surface of Bruch membrane. C. Photomicrograph shows elliptical granules of RPE melanin in apical cytoplasm of RPE cells. The choriocapillaris abuts the outer surface of Bruch membrane. D. Fluorescent microscopy discloses yellow granules of autofluorescent lipofuscin pigment in the basal cytoplasm of the RPE cells. (B. SEM ×1,250, C. Toluidine blue ×250, UV fluorescent microscopy ×400)
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


