Vestibular Function and Anatomy
Christopher J. Holt
Shawn D. Newlands
Nearly all animals have ways of monitoring their own movements and orienting themselves with respect to various sensory cues in their environment. In vertebrates, such behaviors depend heavily on a functioning vestibular system, a critical sensory system that begins in the inner ear as a collection of peripheral detectors for motion and gravity and ends among a number of higher neural centers within the central nervous system (CNS). These higher centers process and integrate incoming motion information from the periphery to inform our perception of our position in space and relative to other objects, to coordinate our movements, and to formulate appropriate reflexes (1). The vestibular system is physically linked to the peripheral auditory system through their cohabitation in the inner ear, and like the auditory system, ultimately converts mechanical forces (i.e., rotation, translation, and/or gravitational forces rather than sound energy) into a neural code that is interpreted centrally. The ability of the vestibular system to detect movements of the head and body resides in the vestibular hair cell whose sensitivity to mechanical stimuli is coupled to a series of electrochemical processes that ultimately change the firing rate of vestibular afferent neurons. Despite the obvious necessity for the vestibular system, we often fail to appreciate its role unless it becomes deficient or defective, at which point, one becomes acutely aware of its importance. In the pages that follow, we discuss the gross anatomy of the peripheral vestibular system, the anatomy and physiology of vestibular hair cells and afferent neurons, the biophysics of sensory transduction, and the organization and functional connectivity of vestibular inputs to the CNS. This chapter serves only as an introduction to the vestibular system; for a more thorough treatment, readers are encouraged to consult dedicated texts.
GROSS ANATOMY OF THE VESTIBULAR SYSTEM
The vestibular labyrinth of vertebrates is an anatomically complex structure consisting of a series of fluid-filled tubes and sacks containing distinct sensory end organs for the detection of both angular and linear acceleration of the head and body (Fig. 143.1). In mammals, the membranous vestibular labyrinth is threaded through a bony labyrinth formed by the dense, hard bone of the otic capsule, within the petrous portion of each temporal bone. A sodium-rich, CSF-like fluid called perilymph bathes the space between the bony and membranous labyrinth, whereas the lumen of the membranous labyrinth is filled with endolymph, a potassium-rich fluid that closely resembles the normal intracellular milieu. Both the perilymphatic and endolymphatic spaces of the vestibular labyrinth are contiguous with the respective spaces of the cochlea. As such, both fluid compartments circulate between the cochlea and the vestibular labyrinth, and thus the homeostatic mechanisms for hearing discussed in Chapter 141 are crucial for vestibular function as well.
The term vestibular labyrinth generally refers to the paired right and left labyrinths together, which are essentially mirror images of each other (see Fig. 143.2A). Each side is comprised of three orthogonally arranged semicircular canals (horizontal or lateral, anterior or superior, and posterior) and two sac-like structures called the utricle and saccule. The utricle and saccule are collectively referred to as the otolith organs. Innervation for the canals, saccule, and utricle all travel in the vestibular nerve portion of cranial nerve VIII. The anterior canal, horizontal canal, utricle, and anterior one-third of the saccule receive their innervation by way of the superior division of the vestibular nerve,
whereas the inferior division innervates the posterior twothirds of the saccule and posterior canal (Fig. 143.1). Upon reaching each end organ, the innervation terminates within the sensory portions of the membranous labyrinth. The sensory neuroepithelium of the semicircular canals and otolithic organs are called the crista and macula, respectively (Fig. 143.1).
whereas the inferior division innervates the posterior twothirds of the saccule and posterior canal (Fig. 143.1). Upon reaching each end organ, the innervation terminates within the sensory portions of the membranous labyrinth. The sensory neuroepithelium of the semicircular canals and otolithic organs are called the crista and macula, respectively (Fig. 143.1).
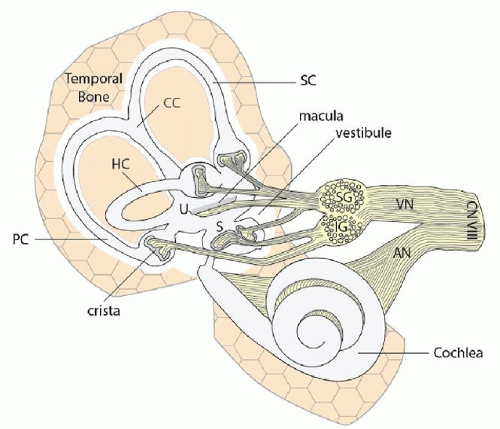 Figure 143.1 The vestibular labyrinth and its innervation. AN, auditory nerve; CC, common crus; CN VIII, cranial nerve 8; HC, horizontal canal; IG, inferior ganglion; PC, posterior vertical semicircular canal; S, saccule; SC, superior or anterior vertical semicircular canal; SG, superior ganglion; U, utricle; VN, vestibular nerve. Shaded areas at each of the nerve endings represent the sensory neuroepithelium. |
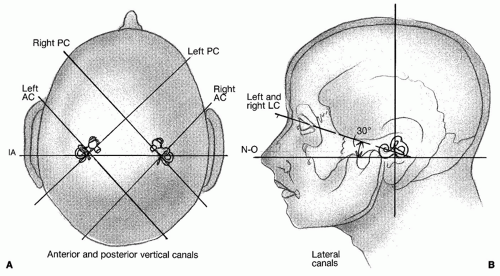 Figure 143.2 A: Orientation of the vertical canals in the human head. B: Plane of the lateral (horizontal) semicircular canals. AC, anterior or superior vertical semicircular canal; IA, interaural; LC, lateral or horizontal semicircular canal; N-O, nasal-occipital axis; PC, posterior vertical semicircular canal. |
One end of each semicircular canal is expanded to form what is called the ampulla. The ampulla is presumably enlarged to accommodate the sensory neuroepithelium and associated accessory structures. The ampulla of either the horizontal or superior semicircular canal is located at the anterior end of each canal, whereas the ampulla of the posterior semicircular canal is located more laterally. The nonampullated ends of the posterior and superior semicircular canals join to form the common crus that subsequently enters the vestibule posteromedially. The nonampullated end of the lateral semicircular canal enters the vestibule posterolaterally.
The semicircular canals detect angular acceleration and thus are well suited for monitoring rotations of the head. A canal’s greatest sensitivity to angular acceleration occurs when the head rotation is made within the plane of that canal. The anatomical organization of the three semicircular canals within the temporal bone is illustrated in Figure 143.2. Horizontal canals are pitched approximately 30 degrees above the horizontal axial plane while the head is upright. The vertical canals (i.e., anterior and posterior) are situated roughly at right angles to the horizontal canals and to each other. Thus, when one looks down at the top of the head, the plane of the anterior canal is oriented at approximately 45 degrees off the midsagittal axis and 45 degrees anterior to the interaural line. The plane of the posterior canal is also 45 degrees off the midsagittal line but aligned roughly 45 degrees behind the interaural line. Thus, the anterior canal on the left is essentially parallel to the posterior canal on the right. The right anterior canal
and the left posterior canal are similarly aligned. The plane parallel to the left anterior canal and right posterior canal passing through the center of the head is perpendicular to the horizontal canal plane and is referred to as the LARP plane. Conversely, the plane perpendicular to both the horizontal canal plane and the LARP plane, but parallel to the right anterior and left posterior plane is referred to as the RALP plane. Because of this canal pairing system (i.e., HC-HC, LA-RP, and RA-LP), movement that excites one canal will inhibit the other, which forms the basis for a push-pull system. As we discuss later, both the excitation and inhibition converge centrally to improve the sensitivity of the system.
and the left posterior canal are similarly aligned. The plane parallel to the left anterior canal and right posterior canal passing through the center of the head is perpendicular to the horizontal canal plane and is referred to as the LARP plane. Conversely, the plane perpendicular to both the horizontal canal plane and the LARP plane, but parallel to the right anterior and left posterior plane is referred to as the RALP plane. Because of this canal pairing system (i.e., HC-HC, LA-RP, and RA-LP), movement that excites one canal will inhibit the other, which forms the basis for a push-pull system. As we discuss later, both the excitation and inhibition converge centrally to improve the sensitivity of the system.
The otolith organs, represented in most mammals and humans by the utricle and saccule, are housed in the major chamber of the vestibular labyrinth called the vestibule. In contrast to the semicircular canals, the otolith organs detect linear acceleration in the plane of each organ. Though neither organ is perfectly planar, the utricular macula is approximately aligned parallel to the earth and is roughly aligned with the ipsilateral horizontal canal (Fig. 143.3). The saccular macula is perpendicular and at right angles to the utricle. Thus, the utricle is sensitive to motion in the horizontal plane, and the saccule is sensitive to movements in the sagittal plane. Collectively, the otolith organs are anatomically organized to detect and monitor linear accelerations along three axes that are perpendicular to each other: interaural, nasal-occipital, and rostral-caudal (Figs. 143.2 and 143.3).
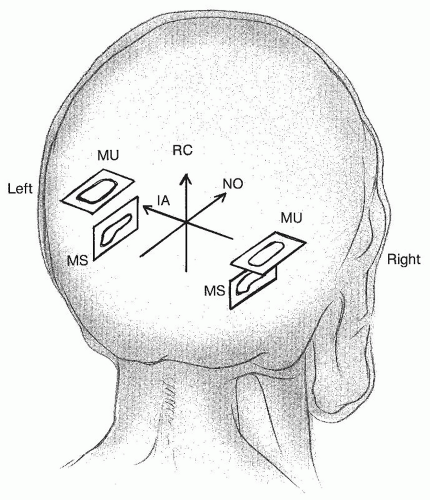 Figure 143.3 Approximate planar layout of the otolith organs. MS, Macular saccule; MU, macular utriculi; IA, interaural axis; NO, nasal-occipital axis; RC, rostral-caudal axis. |
CELLULAR ANATOMY OF THE VESTIBULAR END ORGAN
In order to understand the cellular basis of how various head movements are detected and encoded by the peripheral vestibular system, further examination of the end organ is warranted. An illustration of the sensory neuroepithelium typical of each vestibular end organ is shown in Figure 143.4A. A cross-section through either the crista or macula reveals four distinct cellular constituents: (a) hair cells, (b) supporting cells, (c) afferent nerve fibers and terminals, and (d) efferent nerve fibers and terminals. The roles that each constituent plays in vestibular physiology and function are highlighted below.
Vestibular Hair Cells
As with the auditory system, sensory transduction in the vestibular system is accomplished by a specialized neuroepithelial cell called the hair cell, so named because of the tuft of finger-like projections located along its apical pole (Fig. 143.4A). It is this apical tuft that confers mechanical sensitivity to both auditory and vestibular hair cells. On the basis of their morphology, physiology, and afferent innervation, vestibular hair cells are classified as either type I or type II (2). The type II hair cell is a prototypical, cylindrically shaped hair cell that receives its afferent innervation as a number of bouton terminals along its basolateral edges. In contrast, the type I hair cell is flask-shaped with a rounded base and constricted apical neck, and is innervated by a cup-like, calyx afferent terminal, which almost entirely encapsulates the type I hair cell. Hair cells communicate with their respective afferent neurons by releasing the excitatory neurotransmitter glutamate from specialized presynaptic structures called synaptic ribbons. Type II hair cells are found in the vestibular end organs of all vertebrates while type I hair cells and calyx afferent neurons are restricted to amniotes (i.e., reptiles, birds, and mammals). Likely emphasizing their respective functional roles, type I and type II hair cells and their respective afferent neurons often follow zonal boundaries and thus are not evenly distributed throughout the crista or macula.
Although most of the hair cell and all of its innervation occupy the perilymphatic space, the hair bundle itself is suspended in the endolymphatic compartment. Here, it projects into a gelatinous accessory structure called the cupula in canals or the otolithic membrane in the utricle and saccule. This particular anatomical arrangement is necessary for the proper function of the hair cell and its detection of head movements. The hair bundles of vestibular hair cells are composed of a single eccentrically placed kinocilium and multiple rows of stereocilia (Fig. 143.4). There are typically 20 to 100 stereocilia per hair cell. The kinocilium is the tallest member of the hair bundle and its position at the edge of the apical surface
indicates the orientation of the hair cell. An orderly array of adjacent stereocilia is organized in a descending staircase formation when moving away from the kinocilium. As can be seen in Figure 143.5, this particular geometrical arrangement confers a so-called morphological polarization vector (PV) or axis, which can be identified by drawing a line from the smallest stereocilia through the kinocilium. It turns out that the functional axis of the hair cell is aligned with the morphological axis. It is along this axis that a hair cell is most sensitive to displacement of the bundle. This is made possible by the expression of mechanoelectrical transduction (MET) channels along the tips of the stereocilia that open during hair bundle deflection (3). The mechanosensitivity of MET channels is provided by filamentous tip links that connect MET channels in a lower stereocilium with the membrane of its taller, adjacent neighboring sterocilium. Relative motion of the stereocilia, upon deflecting the hair bundle toward the kinocilium, exerts tension on the tip links thereby prompting the opening of MET channels. What results is an influx of potassium ions from the endolymphatic compartment into the stereocilia that subsequently depolarizes the hair cell. Conversely, deflection of stereocilia away from the kinocilium decreases tip link tension, closes MET channels, and subsequently hyperpolarizes the hair cell. Displacement of the stereocilia perpendicular to the polarization axis, however, causes little to no response in the hair cell. In any one sensory end organ, neighboring hair cells tend to have PVs that are aligned. This is certainly true of canal hair cells whose hair bundles all face the same direction. Hair bundle orientation in otolithic maculae, however, varies considerably particularly along the striola, a line where hair bundle orientation reverses direction nearly 180 degrees (see Fig. 143.6).
indicates the orientation of the hair cell. An orderly array of adjacent stereocilia is organized in a descending staircase formation when moving away from the kinocilium. As can be seen in Figure 143.5, this particular geometrical arrangement confers a so-called morphological polarization vector (PV) or axis, which can be identified by drawing a line from the smallest stereocilia through the kinocilium. It turns out that the functional axis of the hair cell is aligned with the morphological axis. It is along this axis that a hair cell is most sensitive to displacement of the bundle. This is made possible by the expression of mechanoelectrical transduction (MET) channels along the tips of the stereocilia that open during hair bundle deflection (3). The mechanosensitivity of MET channels is provided by filamentous tip links that connect MET channels in a lower stereocilium with the membrane of its taller, adjacent neighboring sterocilium. Relative motion of the stereocilia, upon deflecting the hair bundle toward the kinocilium, exerts tension on the tip links thereby prompting the opening of MET channels. What results is an influx of potassium ions from the endolymphatic compartment into the stereocilia that subsequently depolarizes the hair cell. Conversely, deflection of stereocilia away from the kinocilium decreases tip link tension, closes MET channels, and subsequently hyperpolarizes the hair cell. Displacement of the stereocilia perpendicular to the polarization axis, however, causes little to no response in the hair cell. In any one sensory end organ, neighboring hair cells tend to have PVs that are aligned. This is certainly true of canal hair cells whose hair bundles all face the same direction. Hair bundle orientation in otolithic maculae, however, varies considerably particularly along the striola, a line where hair bundle orientation reverses direction nearly 180 degrees (see Fig. 143.6).
 Figure 143.4 Anatomy and physiology of the sensory neuroepithelium. A: Cellular constituents of the sensory neuroepithelium. BA, bouton afferent; BM, basement membrane; CA, calyx afferent; EN, efferent neuron; HCN, hair cell nucleus; I, type I hair cell; II, type II hair cell; K, kinocilia; S, stereocilia; SC, supporting cell; SR, synaptic ribbon. Hair cells continuously release neurotransmitter that depolarizes afferent terminals to generate action potentials. Recording from a calyx afferent reveals the baseline discharge (bottom). B: Coupling acceleration to hair cell stimulation and afferent discharge. Rotation or translation to the left results in an inertial shift of the cupula or otoconial mass to the right. This shift deflects the hair bundle toward the kinocilium, which opens MET channels in the stereocilia to further depolarize the hair cell. Hair cells subsequently release more glutamate on to afferent terminals resulting in an increase in action potential discharge. Note the increase in the rate of action potentials (bottom). |
Like most neurons, hair cells are able to maintain a negative resting membrane potential, which is necessary for conveying mechanical events at the bundle to vestibular afferent neurons that synapse along their basolateral edges. In the absence of a vestibular stimulus (i.e., no bundle deflection), enough MET channels are thought to remain open to sufficiently depolarize vestibular hair cells. This baseline depolarization permits the continuous release of glutamate from synaptic ribbons located along the basal pole of the hair cell. Once released, glutamate then activates postsynaptic glutamate receptors to depolarize adjacent afferent nerve terminals. Afferent
depolarization results in the generation of action potentials in the afferent neuron that are then transmitted to the brain (Fig. 143.4A



depolarization results in the generation of action potentials in the afferent neuron that are then transmitted to the brain (Fig. 143.4A
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
