(1)
University of Sydney, Sydney, Australia
Overview
1.
Structure and origins
The retinal pigment epithelium (RPE) is a hexagonally packed, monolayer of cuboidal epithelial cells that separates the neural retina from the choroid.
2.
Relationship with neighboring tissues (Fig. 9.1)
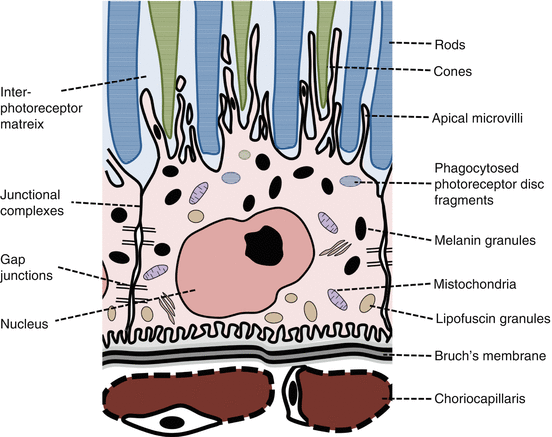
Fig. 9.1
Structure of the retinal pigment epithelial cell
Two specialized extracellular matrices on the RPE basal and apical surfaces enable this interaction:
(i)
Bruch’s membrane
This 5-layered membrane is a molecular sieve that partly regulates the reciprocal exchange of oxygen, fluids, nutrients, and waste products between the retina and choriocapillaris [3].
Rich in elastin and collagen, it provides only a minor contribution to the blood-retinal barrier.
(ii)
Interphotoreceptor matrix
This extracellular matrix is an interface between rod and cone outer segments (OS) and RPE cells [4].
It is found in the subretinal space, consisting of loosely organized proteins and proteoglycans.
3.
Functions of the retinal pigment epithelium (Table 9.1)
Function | |
|---|---|
1 | Photoreceptor outer segment (OS) phagocytosis and renewal |
2 | Light absorption and antioxidant protection |
3 | Vitamin A metabolism and storage |
4 | Barrier function (blood-retinal barrier) and control of fluid and ion transport between the retina and choriocapillaris |
5 | Retinal adhesion |
6 | Photoreceptor alignment |
7 | Secretion of growth factors and immune modulators |
Structure of the Retinal Pigment Epithelium
1.
Gross structure
The RPE extends to the ora serrata where it is continuous with the ciliary pigment epithilium.
It ends posteriorly at the border of the optic disc.
2.
Cellular organization [8, 9]
The RPE contains approximately 3.5 million cells arranged in a regular hexagonal pattern.
At the posterior pole, the cells are tall slender, and densely packed.
Towards the periphery, the cells are flatter, wide, and pleomorphic.
On average, there are 23 photoreceptors per RPE cell [10].
3.
Cell architecture (Fig. 9.1)
RPE cells are polarized with distinctive apical and basal membranes [11].
Anterolaterally, the cells are joined by junctional complexes that contain numerous tight junctions.
This forms an effective barrier for fluid and solutes between the choroid and subretinal spaces [13].
Beneath the junctional complexes, numerous gap junctions link the cells electrically [14].
RPE cells contain numerous apical pigmentary melanin granules that absorb light [14].
Lipofuscin granules, containing residue of digested photoreceptor material, are found basally in the RPE cells. They are more numerous towards the posterior pole and fovea [15].
Functions of the Retinal Pigment Epithelium
RPE cells are highly metabolically active.
1.
Phagocytosis of photoreceptor outer segments (OS) (Fig. 9.2)
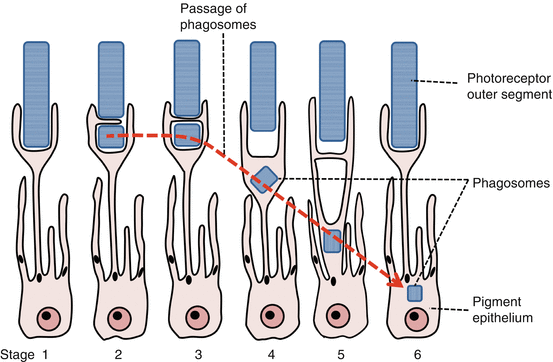
(i)
Phagocytosis allows outer segment renewal.
(ii)
Regulation of phagocytosis.
Outer disc shedding follows circadian regulation and is maximal after morning light onset.
It takes approximately 11 days to renew the whole length of the OS [18].
OS binding is coordinated by the RPE apical receptor α v β 5 -integrin, OS internalization by CD36, and activation of phagocytosis by receptor tyrosinekinase c-mer (MerTK) [19].
(iii)
RPE phagocytic load.
RPE cells have a high phagocytic load, ingesting and degrading much OS material through life.
This phagocytic and metabolic load causes RPE lipofuscin accumulation with age [20].
2.
Light absorption and anti-oxidative protections
(i)
Light absorption leads to heat generation.
(ii)
Retinal pigment epithelial cell oxidative stress
Excess light, heat, and O2 expose RPE cells to oxidative damage [8].
High metabolic activity and age-related lipofuscin accumulation exacerbate oxidative stress.
3.
Vitamin A metabolism and the visual cycle (Fig. 9.3)
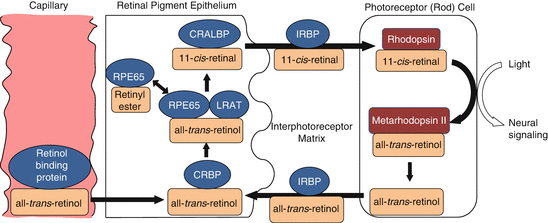
RPE is involved in the storage and metabolism of vitamin A (retinol) and its derivatives (retinoids).
(i)
(ii)
RPE storage and activation of vitamin A
99% of RPE vitamin A is stored in cytplasmic droplets as retinyl ester, a stable, nontoxic form [26].
This can be converted to 11-cis-retinal, the key chromophore of the visual pigments.
This conversion occurs via a complex involving RPE65, which acts as an isomerase, and lecithin:retinol transferase (LRAT) [27, 28].
11-cis-retinal binds to cellular retinol-binding protein (CRALBP) within the RPE.
(iii)
Vitamin A transport to the photoreceptor OS
11-cis-retinal is shuttled across the subretinal space by interphotoreceptor matrix retinal binding protein (IRBP) [ 29].
The chromophore 11-cis-retinal forms a complex with a protein (opsin) to form visual pigment (rhodopsin in rods) within OS discs.
(iv)
Light reaction and subsequent events
Light induces a conformational change of the chromophore from 11-cis-retinal to all-trans-retinal.
All-trans-retinal leaves the disc membrane via ATP-binding cassette protein transporter ABCR4.
It is converted into all-trans-retinol and transported back to the RPE via IRBP [31].
Within the RPE cell, it binds to cellular retinol-binding protein (CRBP) and interacts with the RPE65/LRAT complex.
(v)
Light adaptation (See Chap. 21. Luminance Range for Vision)
Vitamin A metabolism is essential in regeneration of photopigments after strong light esposure [33].
In light, there is a rapid turnover of retinal; in the dark, turnover occurs more slowly.
Rapid bleaching of pigment in light and slower photopigment regeneration in dark are important components of visual adaptation to different light intensities.
Pigment regeneration involves sequential recruitment of vitamin A sources IRPB, CRALBP, and RPE65 [34].
4.
Barrier function and fluid and solute transport (Fig. 9.4)
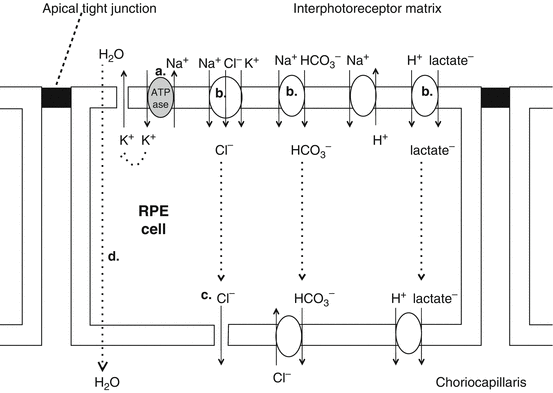
Fig. 9.4
Solute and fluid movement across RPE cells
(i)
Blood-retinal barrier maintenance (See Chap. 11. Ocular Circulation)
The tight junctional complexes around the RPE cells maintain the blood-retinal barrier (the barrier between blood from the choroid and OS of the photoreceptors) [35].
This regulates solute flow to the retina, maintaining tight control of extracellular composition.
The blood-retinal barrier also maintains the immune privilege of the eye [36].
(ii)
Transepithelial transport
Due to high paracellular resistance due to intercellular tight junctions, molecules and ions flow across the RPE via transepithelial transport to:
a.
Supply nutrients to the photoreceptors
b.
Control ion homeostasis
(iii)
Metabolic pump
(b)
(d)
This energy-dependent transfer of solutes and water provides the RPE with a capacity for pumping out excess fluid despite high oncotic pressure of the interphotoreceptor matrix [44].
(iv)
Oxygen supply
The choriocapillaris is the main source of oxygen for the outer retina [45].
Oxygen freely diffuses across Bruch’s membrane and the RPE to supply the outer retina.
5.
Retinal adhesion
Get Clinical Tree app for offline access

Possible mechanisms of retinal adhesion to the RPE include:
(b)
Interdigitation of RPE apical villous processes with photoreceptor outer segments [12].
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree

