(1)
University of Sydney, Sydney, Australia
Overview
The primary visual cortex (V1) receives visual information from segregated magnocellular, parvocellular, and koniocellular channels of the lateral geniculate nucleus (LGN) [1, 2].
Separation of these channels is largely preserved in V1 processing [3].
V1 also receives modulatory input from several non-LGN cortical and subcortical areas.
V1 sends these basic image descriptors to extrastriate visual association areas 2, 3, 4, and 5 (V2, V3, V4, and V5) for higher visual analysis of specific stimulus attributes (see Chap. 15, The Extrastriate Cortex) [7].
Structure of V1
1.
Anatomy (Fig. 14.1)
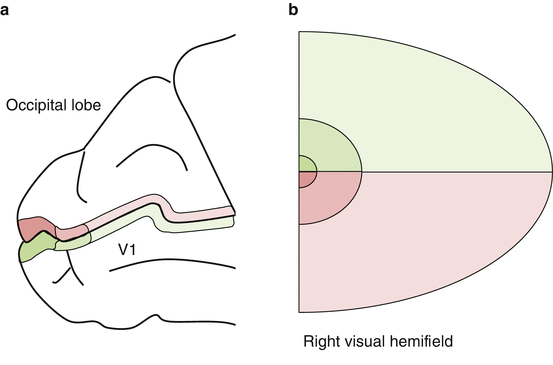
Fig. 14.1
(a) Location of V1 within the occipital lobe, with (b) visuotopic projection of the hemifield
V1 is located within the occipital lobe of the cerebrum [10].
It extends along the medial wall from the posterior pole on either side of the calcarine sulcus.
Like all cerebral cortex, it contains six principal layers.
V1 is called the striate cortex due to a heavily myelinated stripe in layer 4, the stria of Gennari [11].
Layer 4 is heavy myelinated due to high density of LGN projections.
2.
Visuotopic organization
V1 in each hemisphere represents the contralateral visual hemifield [12].
Each side receives uncrossed ipsilateral and crossed contralateral fibers from the LGN.
In V1 the peripheral field is represented anteriorly and the central field posteriorly.
3.
Layers of V1 (Fig. 14.2)
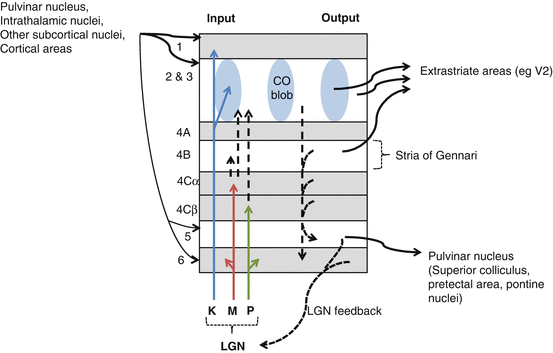
Fig. 14.2
V1 layers and connections
An overview of the connections of the layers is outlined in Table 14.2 and Fig. 14.2.
Layer
Input
Output
1
Modulatory subcortical input
Koniocellular LGN input
Other V1 layers
2 & 3a
Projections from layer 4
Extrastriate areas
4
Most LGN inputs
Layers 3, 5, and 6
5
Projections from layer 4
Subcortical areas (thalamus, pons, and midbrain)
6
Projections from layer 4
Subcortical areas (thalamus, pons, and midbrain)
4.
Cytochrome oxidase blobs
Layers 2 and 3 contain areas that stain for the mitochondrial enzyme cytochrome oxidase (CO), known as the “CO blobs.”
CO blob neurons have color-opponent receptive fields and lack orientation selectivity [23].
Connections of V1 (Fig. 14.2)
1
Inputs to V1
(i)
Lateral geniculate nucleus inputs
LGN inputs, predominantly into sublayer 4C, determine the activation of V1 neurons.
The inputs are segregated according to:
This segregation is maintained at the first synapse in V1.
M cells terminate in layer 4Cα which projects to sublayer 4B; P cells terminate in layer 4Cβ which projects to sublayer 4A [1, 2, 5, 25, 27].
Sublayer 4A receives collateral input from P and K cells and projections from cells in 4B.
Sublayer 4A may have a role in synthesizing information from separate M, P, and K streams [28].
Neurons in layer 4 send off connecting axons principally to layer 3 and also to layers 5 and 6.
(ii)
Non-lateral geniculate nucleus inputs
V1 receives modulatory inputs from cortical and subcortical areas.
These regulate the signals sent from V1 to higher-order areas for further processing.
Subcortical inputs include those from the thalamic intralaminar and pulvinar nuclei (which synapse in V1 layer 1), the amygdala, and the basal forebrain nuclei [29–32].
2
Output pathways for V1
(i)
Output from layer 3
Layer 3 provides output to a number of extrastriate visual cortical areas (V2, V3, V4, and V5) [21].
(ii)
Binocularity and Ocular Dominance Columns
1.
Ocular dominance columns
Input arriving from the LGN into layer 4C remains segregated according to the right or left eye [39].
Subsequent connections to binocular cells in layers 3, 5, and 6 combine input from both eyes [40].
Binocular neurons tend to display a preference for one eye and are organized into ocular dominance columns (ODCs) (Fig. 14.3) [39, 41].
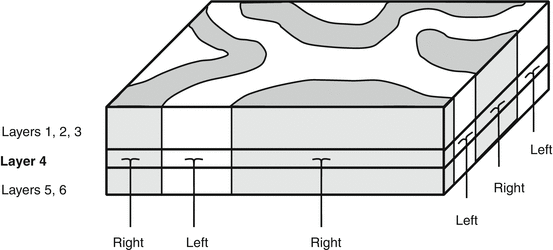
Fig. 14.3
Ocular dominance columns
The ODC extends from layers 1 to 6 reflecting the laterality of layer 4C cells within that column [42].
ODCs are organized into adjacent, alternating bands of V1.
In V1 each point in visual space is represented by two ODCs, one for each eye.
2.
Development of ocular dominance columns
Cortical visual development occurs after birth in response to visual stimuli.
3.
Visual deprivation
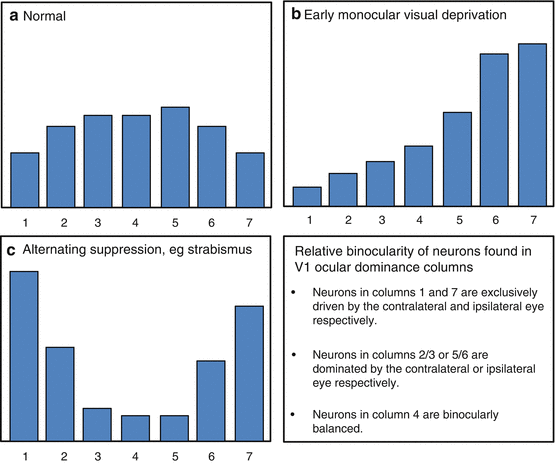
Receptive Field Properties of V1 Cells
V1 cells transform the visual signal from LGN cells.
1.
Orientation sensitivity
(i)
Simple cells
Simple cells have elongated center-surround receptive fields aligned at a particular orientation [49].
These receptive fields are formed by the summation of overlapping LGN cell fields (Fig. 14.5).
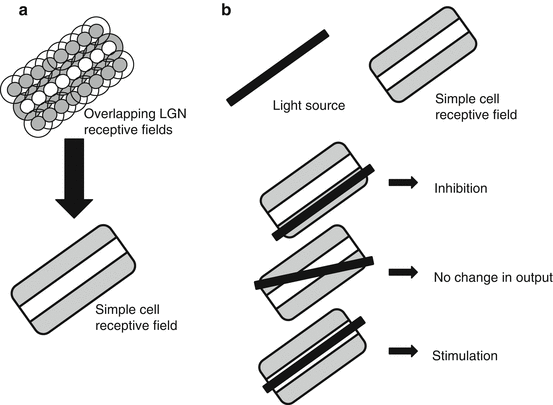
Fig. 14.5
Simple cell receptive fields. (a) Summation of LGN receptive fields; (b) Stimulus responses
Simple cells respond to a line of light at a specific orientation.
Their receptive fields vary according to orientation and length [50].
(ii)
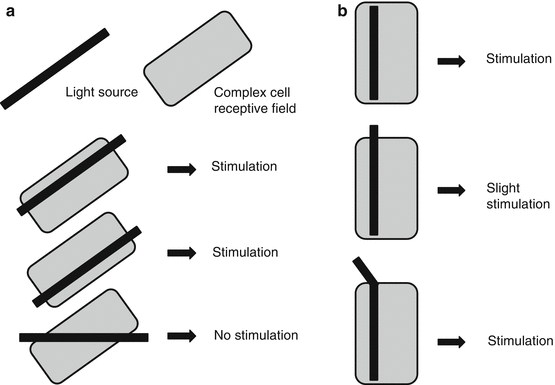
(iii)
End-stopped cells (Fig. 14.6b)
These respond only if a correctly oriented stimulus is of appropriate length.
If the stimulus extends beyond the receptive field, the response is diminished, but not if the extending part of the stimulus has a different orientation to the receptive field.
2.
Motion sensitivity
Motion is coded by certain V1 neurons.
These are sensitive to temporal delay between two adjacent stimuli with the same orientation [58].
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


