(1)
University of Sydney, Sydney, Australia
The Lens
Overview
1.
Structure
The lens is a biconvex, transparent structure located behind the iris (Fig. 4.1).
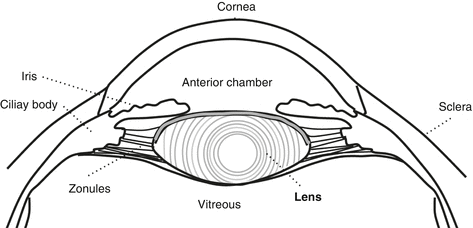
Fig. 4.1
The lens and surrounding structures
It consists of:
(a)
An elastic lens capsule
(b)
An anterior single layer of cuboidal epithelial cells
(c)
Elongated lens fiber cells
2.
Refractive power
The lens provides 15 diopters of the total optical power of the eye.
It is capable of varying that power on accommodation, allowing the eye to vary its focal point.
This permits a clear retinal image for objects that are either distant or near.
Development (Fig. 4.2) [3]
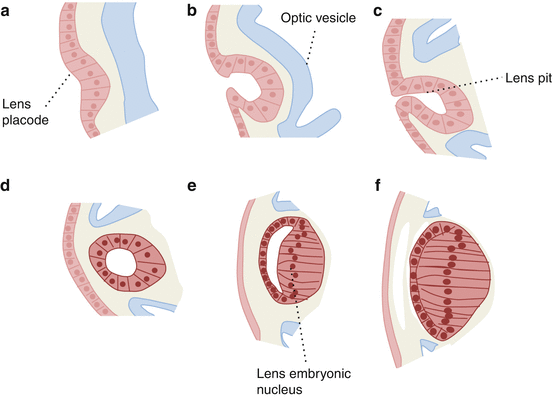
Fig. 4.2
Development of the lens
At 4 weeks gestation the optic vesicle, an outgrowth of the forebrain, makes contact with the surface ectoderm inducing a localized thickening, the lens placode (a) [4].
The lens pit then forms by invagination (b, c).
The lens vesicle separates from the surface at 5–6 weeks (d).
Posterior lens vesicle cells elongate, lose their nuclei, and become the embryonic nucleus (e–f).
From week 6 the lens capsule develops, while the lens is enveloped by a delicate vascular system, the tunica vasculosa lentis.
By week 12 the tunica vasculosa becomes more extensive and is supplied by the hyaloid artery.
At 7 months, the vascular system regresses.
Optical Properties
1.
Refractive properties [5–7]
The refractive properties of the lens are due to:
(a)
Crystallin proteins in fiber cells at high concentration with a higher refractive index than aqueous (Table 4.1)
Media
Surface
Radius of curvature (mm)
Refractive index
Refractive power (diopters)
Air
1
Tear film
Anterior surface
7.7
1.336
+43.6
Cornea
Anterior surface
7.8
1.376
+5.3
Posterior surface
6.9
−5.8
Aqueous fluid
1.336
Lens
Anterior surface
11.0
1.362–1.406 (periphery – center)
Approx +15.0
Posterior surface
−6.5
(b)
The radii of curvature of the lens refractive surfaces
2.
Transparency
Normal transparency is maintained by:
(i)
The regular arrangement of lens fibers
This depends on newly formed lens fibers that mesh precisely with the older, underlying fibers [2].
(ii)
The homogenous, non-particulate lens fiber cytoplasm [9]
As fibers mature, intracellular organelles degenerate, leaving crystallins dominant in the cytoplasm.
(iii)
A highly reducing biochemical environment
This counteracts oxidative stress caused by molecular oxygen or free radicals [10].
(iv)
The paracrystalline state of proteins
This allows a supersaturation of crystallins without formation of aggregates [11].
Structure
1.
2.
Lens capsule
The lens capsule is an elastic basement membrane that envelopes the whole lens.
It is composed of type IV collagen in a matrix of glycoproteins and sulfated glycosaminoglycans [12].
It is synthesized primarily by epithelial and superficial fiber cells; posteriorly, capsule growth virtually ceases early in life but continues anteriorly with age [13, 14].
It is thickest in the anterior midperiphery (21 μm). At the anterior pole it is 13 μm; it is thinnest at the posterior pole (4 μm) [8].
The dense outer layer (zonular lamella) receives zonular insertions [15].
These zonules exert tension which produces flattening of the lens in the unaccommodated state.
3.
Epithelium
The epithelium is a monolayer of cuboidal cells inside the anterior and equatorial capsule.
The epithelial cell lateral membrane interdigitates with adjacent cells, adjoined by:
(a)
Desmosomes for cell adhesion
Epithelial cells have plentiful Na + /K + ATPase metabolic pumps and many other proteins important for lens metabolism and ion transport. [17]
They are responsible for:
Toward the equator, in the germinative zone, cells are higher, more narrow and cylindrical.
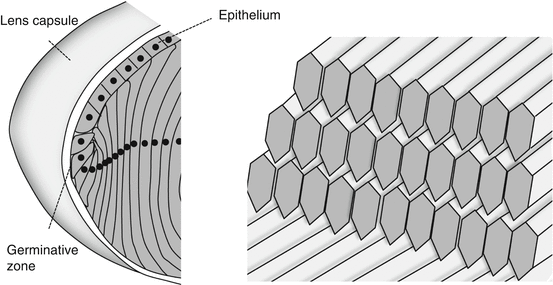
4.
Lens fibers [2]
Lens fiber cells are elongated, densely packed cells supersaturated with crystallins:
On differentiation they become hexagonal in cross section and lose their organelles (Fig. 4.3b) [9, 19, 20].
In the cortex they are 8–12 mm long, 10 um wide and 4.5 um thick. Intercellular distance is 20 nm [21].
Fibers meet at the anterior and posterior polar sutures, which are Y shaped in the embryonic lens [22].
Mature fibers are buried deep in the center by accumulation of successive fiber layers; peripheral fibers are newer.
In this way the lens increases in size and fiber number through life [23].
Deeper lens fibers have ball and socket interlocking regions. These prevent slippage during changes in lens shape [24].
Abundant gap junctions between the cells (the highest concentration in the body) allow rapid intercellular movement of small molecules and ions [16].
5.
Zonules
The zonules are fine fibrillary structures that suspend the lens from the ciliary body (Fig. 4.1):
They are composed of fibrillin, elastin, mucopolysaccharide and glycoproteins [25].
They extend from the pars plana toward the pars plicata passing between the ciliary processes.
The zonules insert onto the lens equatorial capsule [15].
They maintain lens stability and exert tonic stretch that relaxes with accommodation.
Lens Proteins
Lens fibers have a low water content and a high concentration of proteins.
This is essential for transparency, a high refractive index and deformability during accommodation.
1.
Lens proteins: crystallins
The predominant proteins of the lens are classed as crystallins.
Crystallins make up 40 % of the wet weight of lens fibers [11].
Their concentration is three times greater than normal protein concentration of most human cells.
(i)
Alpha crystallin (30 % of total lens protein) [26]
These are large proteins (7 × 105 daltons (Da)) that form complexes composed of αA or αB subunits.
Functions include:
α-crystallin has auto-kinase activity; the role of this in lens metabolism is not yet determined [29].
With age, these aggregate into large insoluble proteins that contribute to loss of lens transparency [30].
(ii)
Beta/gamma crystallin (56 % of total lens protein) [31]
Initially thought to be distinct protein families, these are now considered one superfamily [32].
β-crystallins exist as large polymers (4 × 104 – 2.5 × 105 Da); γ-crystallins exist as monomers.
β-crystallins and γ-crystallins can bind Ca2+ and may have a role in cytoplasmic calcium buffering [31].
γ-crystallin is implicated in cold cataract and precipitates on cooling (<10 °C); on rewarming this reverses [33].
2.
Non-crystallin proteins
These are predominantly structural proteins and metabolic enzymes, including:
(i)
(ii)
(iii)
Enzymes
For example, transport enzymes, ATPase, alkaline phosphatase, adenyl cyclase, and dehydrogenases
Lens Electrolytes and Metabolism
The electrolyte composition of lens as a whole resembles a single cell [39].
Relative to the surrounding aqueous and vitreous, the lens has:
(a)
High K + (125 mmol/L)
(b)
Low Na + (14 mmol/L)
(c)
Low Cl − (26 mmol/L)
The high prevalence of cell-to-cell gap junctions for rapid exchange between cells allows the lens to function as a syncytium (like a single cell) [16].
1.
Sodium (Fig. 4.4a)
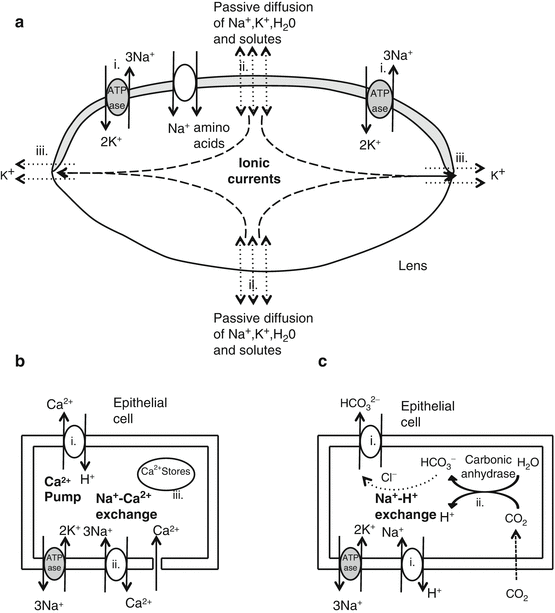
Fig. 4.4
Models for ionic fluxes in the lens. (a) Sodium flux, (b) calcium flux, and (c) pH regulation
(i)
The epithelium has active Na + /K + ATPase pumps to extrude Na+ and accumulate K+ in the lens [40].
This ionic gradient provides the energy for other processes including:
(a)
Na+/Ca2+ exchange
(b)
Na+/HCO3 2− co-transport
(c)
Amino acid transport
These help maintain high intracellular levels of HCO3 2− and amino acids.
4.
Carbohydrate metabolism [4, 46]
Glucose is the principal carbohydrate of the lens.
It enters the lens from the aqueous by simple diffusion and insulin-dependent facilitated transfer.
Anaerobic metabolism is highly prevalent in the lens, compared to most body tissues, because of:
(a)
Low oxygen tension.
(b)
Lens fibers lack mitochondria necessary for aerobic metabolism.
Compared to aerobic metabolism, anaerobic glycolysis:
(a)
Produces fewer free radicals
(b)
Requires little oxygen, both of which help maintain lens transparency
Metabolic pathways in lens glucose metabolism are listed below (Table 4.3, Fig. 4.5):
Table 4.3
Metabolic pathways in lens glucose metabolism
Metabolic pathway
Glucose utilized (% of total)
ATP produced (% of total)
Anaerobic glycolysis
80
66
Aerobic respiration
3
20
Hexose monophosphate shunt
15
Sorbitol pathway
5
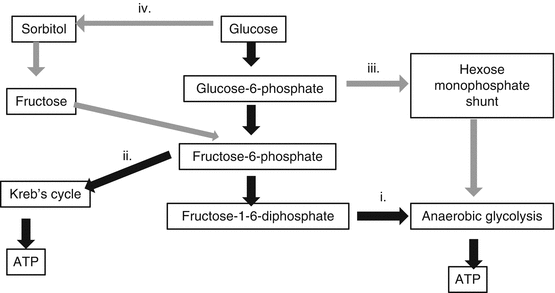
Fig. 4.5
Carbohydrate metabolic pathways used by the lens
(i)
Anaerobic glycolysis
Anaerobic glycolysis produces lactic acid that is partly used for the Kreb’s cycle but mostly diffuses into the aqueous.
This causes a high aqueous concentration of lactate.
(ii)
Aerobic respiration (including Kreb’s or tricarboxylic acid cycle)
Aerobic respiration occurs in the epithelium where the necessary O2 and enzymes are available [48].
These produce CO2 which diffuses into the aqueous.
(iii)
Hexose monophosphate shunt
NADPH is a cofactor in many biochemical reactions including the maintenance of reduced glutathione by glutathione reductase.
(iv)
Sorbitol pathway [50]
Glucose is converted to sorbitol and then fructose via aldose reductase and polyol dehydrogenase.
This pathway is possibly a means of protecting the lens from osmotic stress in hyperglycemia.
5.
Lipids
Get Clinical Tree app for offline access

(i)
Cholesterol and sphingomyelin
The lens fiber membrane has a unique lipid composition, with high cholesterol and sphingomyelin levels; the cholesterol levels are the highest of any cell types in the body [51].
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree

