(1)
Department of Ophthalmology and Visual Sciences, University of Iowa Hospitals and Clinics, Iowa City, IA, USA
The subject of the blood-retinal barrier (BRB) is important in understanding the pathology of various retinal diseases as well as in their treatment. There is a voluminous literature on the BRB and its derangement. The retina is transparent so that the light travels unhindered to the photoreceptors. Therefore, for proper visual function of the retina, as well as for physiological integrity of the retinal vessels and retinal homeostasis, the presence of an efficient BRB is essential. To do that, the BRB regulates movement of fluids and molecules between the blood and the retina. Also for drug delivery to the retina, the presence of a BRB becomes an important consideration [1].
The BRB is located at two places: (1) An inner BRB is due to complex tight cell junctions (zonulae occludentes) of retinal capillary endothelial cells. (2) An outer BRB is due to tight cell junctions in the retinal pigment epithelium (RPE) layer. These tight cell junctions prevent diffusion of substances and exercise a high degree of control on permeability of solute and fluid between the ocular vascular bed and neural retinal tissues. The BRB is of great clinical importance because breakdown of it results in a variety of retinal pathologies; for example, derangement of the inner BRB results in macular edema and of the outer BRB in serous retinal detachment.
Inner Blood-Retinal Barrier
This was described by Cunha-Vaz and colleagues [2–4]. It is primarily due to complex tight cell junctions (zonulae occludentes) of retinal capillary endothelial cells. Retinal astrocytes, Müller cells, and pericytes also contribute to the proper functioning of the inner BRB [5], and they may influence tight cell junction formation and maturation [6]. Let us look at the role of the various components of the inner BRB:
Endothelial Cells: A tight cell junction among them is the main constituent of this BRB.
Müller Cells: They extend their endfeet onto blood vessels of the retina and cover them [7], suggesting a close spatial relationship between Müller cells and blood vessels. They induce the formation of barrier properties by vascular endothelial cells. Thus, they play a major role in the formation of barrier properties in retinal vessels [8]. They also play a role in the uptake of nutrients and the disposal of metabolites [9, 10]. Müller glial cell-derived factors modulate endothelial cell functions [11].
Glial Cells: Astrocytes extend their endfeet onto the wall of retinal blood vessels and sheath them [7] and thereby influence the barrier function [12]. They enhance BRB properties, at least in part by increasing tight junction protein expression [13]. Also glial cells play an important role in vessel integrity and barrier properties, both via direct contact [14] and through release of humoral factors [15]. In particular, glial cell line-derived neurotrophic factor has been recognized to enhance barrier tightness [16], whereas transforming growth factor beta appears to decrease it [17]. Glial cell-derived cytokines, such as glial cell line-derived neurotrophic factor, neurturin, and vascular endothelial growth factor, regulate BRB function [16, 18]. Degeneration of astrocytes in feline retinopathy of prematurity causes failure of the blood-retinal barrier [19].
Pericytes: There is a high density of pericytes in the retina as compared to other organs, including the brain [20–23]. Pericytes interact with endothelial cells to form a fully functional BRB [24]. Pericytes have contractile proteins, in particular smooth muscle actin, desmin, and non-muscle myosin [24–26]. The myogenic mechanism for vascular autoregulation of blood flow is due to both pericytes and smooth muscle cells. They are able to regulate the capillary diameter through contraction and relaxation [27–29]. In addition, pericytes have a variety of receptors for vasoactive substances, enabling them to respond to endothelin-1 by contraction [30] and to nitric oxide by relaxation [31], suggesting an intimate relationship between pericytes, the endothelium, and also the surrounding glia in regulating the functional dynamics of the vascular wall.
In summary, the tight junction of endothelial cells in the inner BRB could be developed by cellular interactions between Müller cells, pericytes, astrocytes, and endothelial cells [32].
The inner BRB is compromised in several conditions, including diabetes mellitus, retinal vein occlusion, and various inflammatory diseases.
Role of Vascular Endothelial Growth Factor (VEGF) in BRB
A large number of studies have shown that VEGF plays an important role in the breakdown of the inner BRB and development of macular edema and various other retinal pathologies. Increased vascular permeability for plasma proteins induced by VEGF in BRB endothelium is predominantly caused by a mechanism involving active trans-endothelial transport via pinocytotic vesicles and not by formation of endothelial fenestrations or vesiculo-vacuolar organelles [35]. Early BRB breakdown in experimental diabetes is VEGF dependent and is restricted, in part, to the venules and capillaries of the superficial inner retinal vasculature [36]. In addition to VEGF, proinflammatory cytokines, nitric oxide, oxidative stress, and inflammation contribute to the loss of the BRB [4, 37]. Glial dysfunction is a primary contributor to the BRB breakdown in retinal vascular diseases [38].
Outer Blood-Retinal Barrier
The outer BRB is located in the RPE, which is a single cell layer and separates the outer surface of the retina from the choriocapillaris (Fig. 9.1). The outer retina is supplied by the choriocapillaris. The RPE has three important functions. (1) Because of the presence of the BRB in the RPE, it regulates the entry of nutrients from the blood in the choriocapillaris to the photoreceptors; (2) it removes waste products; and (3) it maintains its adhesion with the retina. Choriocapillaris have fenestrations, so that they leak profusely, allowing passage of large molecules such as albumin into the extravascular spaces of the choroid. The tight cell junctions of the RPE, which constitutes the outer BRB, prevent diffusion of fluid from the choroid into the retina. Initially during development, RPE cell junctions are leaky, and later on, they develop tight cell junctions [43]. GTPase Rap1 regulates the formation and maintenance of the RPE cell junctional barrier; it is an important regulator of RPE cell junctions and is required for maintenance of barrier function [44].
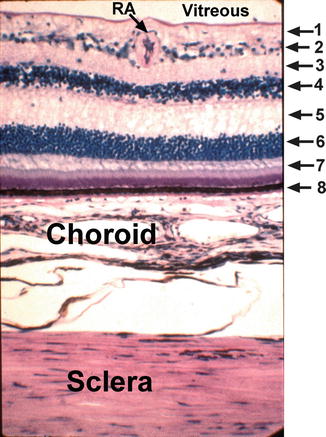
Fig. 9.1
Light micrograph of the retina, choroid, and sclera. The retinal layers are identified by the following numbers: 1 nerve fiber layer, 2 ganglion cell layer, 3 inner plexiform layer, 4 inner nuclear layer, 5 outer plexiform layer, 6 outer nuclear layer, 7 rod and cone layer, 8 retinal pigment epithelial layer, RA retinal arteriole (Reproduced from Hayreh [42])
Oxidized low-density lipoprotein could promote senescence of RPE cells, which leads to outer BRB dysfunction as an early pathogenesis of age-related macular degeneration [45]. In hypertensive choroidopathy, because of ischemia of the RPE, there is breakdown of the outer BRB resulting in serous retinal detachment [46]. Breakdown of the outer BRB has also been reported in diabetes and ischemia [47].
For transport across the RPE, these cells show marked apico-basal polarity, with long apical microvilli and deep basal infoldings [48]. Nutrients and vitamin A are transported in baso-apical direction from the blood to the photoreceptors. Removal of metabolites, extracellular fluid, and ions takes place by transcellular transport from the subretinal space toward the choriocapillaris bed.
Polarity of the Outer and Inner BRBs
It is the protein complex at the tight cell junction that allows the establishment of the polarities of the BRB, restricting paracellular diffusion of blood barrier compounds into the neuronal tissues [3, 49]. Creation of cell polarity is a distinguishing feature of a tissue barrier. This cell polarity is associated with organization of the cytoskeleton, apical/basal cell membrane proteins, and organization of the junctional complexes between neighboring cells [50]. The ability of RPE to regulate transport through it depends upon two properties: (1) presence of tight cell junctions to prevent diffusion through the spaces in between the cells and (2) an asymmetric distribution of proteins to regulate vectorial transport across the monolayer [43].
There is no barrier within the retinal tissue itself [46], so there is a constant movement of small molecules (mainly water) from the vitreous cavity into the inner retina and through the RPE to the choroid. The internal limiting membrane prevents diffusion of macromolecules but allows the passage of smaller molecules.
Functions of the BRB
The retina is a transparent structure, and that is essential for its normal optical visual function, for the light to go uninterrupted to the photoreceptors. For proper physiological integrity of the retinal vessels, microenvironment, homeostasis, and visual function of the retina, an efficient BRB is essential. The tight interendothelial cell junctions block movement of macromolecules toward the retinal interstitial space. A collection of fluid in the retina not only causes edema but also may interfere with its transparency.
The transport of nutrients to the retina and removal of waste products from it through the BRB occurs in two ways: (1) transcellular and (2) paracellular. By the transcellular pathway, most proteins are transported nonselectively within vesicles, either in their fluid phase or adsorbed to the vesicular membrane [51]. Constitutive transcytosis of albumin across the vascular endothelium is done by adsorption or as bulk fluid and is implicated in regulating the transvascular oncotic pressure gradient of albumin [52, 53]. By the paracellular pathway, generally water, ions, and small non-charged solutes are transported by passive diffusion and with low selectivity along electrochemical and osmotic gradients that are built up by the activity of transcellular transporters or by external gradients of solutes [54].
The outer retinal layers, including the photoreceptors, and peripheral retina are avascular. They are supplied by the choroidal vascular bed. RPE regulates entry of nutrients from the blood in the choriocapillaris to the photoreceptors and also eliminates waste products. Tight cell junctions between the RPE cells prevent leakage of fluid from the choroid to the retina, helping to maintain RPE adhesion with the retina. This barrier breaks down when the RPE cells are destroyed or subjected to ischemia, as in hypertensive choroidopathy (Fig. 9.2) [46].
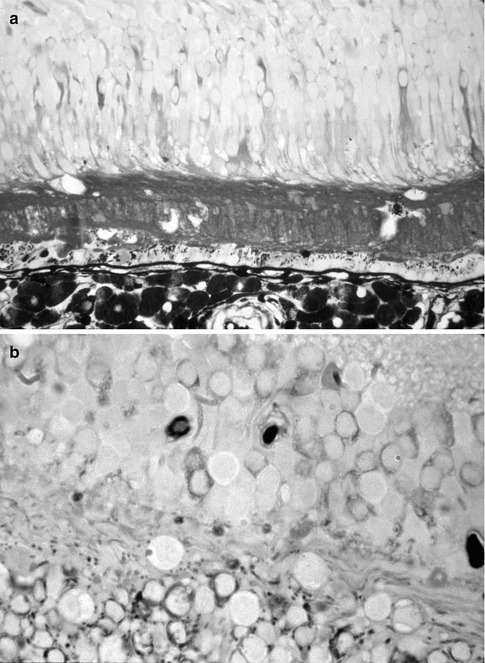
Fig. 9.2
Histological sections of an eye with hypertensive choroidopathy and with serous retinal detachment and macular edema, after horseradish peroxidase (HRP) intravenous injection. Notice the leakage of HRP (brown) from the choriocapillaris into the subretinal fluid (in a) and into the retinal tissue (in a, b) (Reproduced from Hayreh et al. [46])
Cunha-Vaz [55] described the transport of major electrolytes in the BRB, with a net K+ flux from retina to blood and a net Mg++ flux in the opposite direction across the BRB sites, and that provides a low K+ and high Mg++ extracellular environment in the retina. There is also absorptive transport of amino acids across the endothelial and chorioretinal components of the BRB, and that creates a continuous “flow” of these important precursors into the retina. Active absorptive transport across the blood-ocular barrier system removes several substances, including organic anions from the extracellular fluid of the retina. He stated that both the outer and inner components of the BRB have a variety of absorptive transport processes, which are capable of removing potentially harmful substances from the extracellular fluid of the retina.
Corticosteroid Therapy and the BRB
Studies have shown that corticosteroids, such as intravitreal triamcinolone acetonide, inhibit breakdown of the BRB.
Mechanisms by Which Corticosteroids Block Blood-Ocular Barrier Breakdown
The following mechanisms have been reported:
1.
2.
Corticosteroids inhibit VEGF-induced vascular leakage by modulating signals or effector proteins downstream of the VEGF receptor [56].
3.
Triamcinolone acetonide has the potential to influence cellular permeability, including the barrier function of the RPE in age-related macular degeneration-affected retinas [60].
4.
Dexamethasone may inhibit retinal accumulation and leukostasis accumulation and vascular permeability through its blockage on VEGF and ICAM-1 expression [58].
5.
Triamcinolone acetonide stabilizes the BRB in association with regulation of VEGF-A, Flk-1, and Flt-1 expression in retinas in the early stages of diabetes [59].
Acetazolamide Therapy in BRB
A fluorescein angiography study showed that acetazolamide increased unidirectional passive permeability by stimulation of the outward active transport of ion and fluid from the retina to the choroid [61, 62], and the edema-reducing effect of acetazolamide is due to decreased leakage and stimulated active transport across the BRB [41].
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


