Synaptic ribbons are electron-dense structures that are located at the base of hair cells and play a role as a pool of neurotransmitters [2]. From the point of view of auditory function, the synaptic contacts between the inner hair cell and type I afferent nerve fiber are particularly important (Fig. 7.1). Synaptic ribbons control release of neurotransmitters at various modes, rapid release of a large amount of neurotransmitters or slow and consistent release of a small amount of neurotransmitters depending on the intensity and duration of sound stimuli [2]. Synaptic ribbons at the base of the inner hair cells face to a single postsynaptic density at the postsynaptic membrane of type I afferent nerve terminal, of which contains large numbers of AMPA-type glutamate receptors [6]. A large number of glutamate receptors may contribute to secure from receptor saturation. Glutamate released from ribbon synapses to synaptic clefts is removed by amino acid transporters including the glutamate/aspartate transport GLAST in supporting cells [7].
7.3 Vestibule
The vestibular sensory epithelium also has two types of hair cells, type I and II hair cells. Type I and II hair cells differ in their afferent innervation patterns (Fig. 7.2). Type I hair cells are surrounded by the calyceal afferent nerve endings, and type II hair cells are innervated by bouton-type afferent nerve endings [8]. Type I hair cells have a single calyceal terminal, while type II hair cells have a number of bouton-type nerve endings. Also in the vestibular system, synaptic transmission between hair cells and afferent nerve fibers is mediated by ribbon synapses. The expression of AMPA-type glutamate receptors was also reported in the postsynaptic dense in afferent nerve endings in the vestibular epithelium [9]. Comparing with cochlear synapses, details of synaptic functions in vestibular hair cells have not been elucidated. Unlike mammalian cochleae, mammalian vestibular hair cells have the potential of spontaneous regeneration [10, 11]. In addition, the capacity of hair cell regeneration by gene transfer has extensively been investigated [12]. The majority of regenerated hair cells exhibited type II morphology.
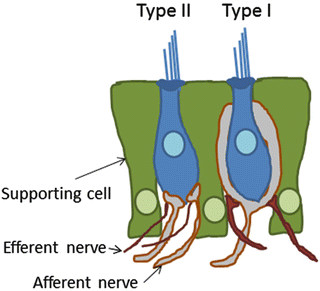
Fig. 7.2
Hair cell synapses in the vestibule
7.4 Synaptic Ribbons and Disease
The dysfunction of synaptic ribbons in inner hair cells of cochleae causes hearing loss. Hearing impairment due to dysfunction of ribbon synapses showed threshold elevation of auditory brain stem response (ABR) thresholds and normal otoacoustic emissions [2]. Based on this, dysfunction of ribbon synapses is considered one of the causes for auditory neuropathy [13]. In animal experiments, the swelling of afferent dendrites of spiral ganglion neurons has well been known as one of the early events that occur in the cochlea after noise damage or exposure to excessive excitatory amino acids [14, 15]. In general, the swelling of afferent dendrites disappears within 7–14 days after the onset of noise- or drug-induced hearing loss. Therefore, this temporal degeneration at the afferent dendrites has been considered one of the causes for temporal thresholds shifts that are observed after noise exposure. In the process of hearing recovery from temporal threshold shifts, regeneration of ribbon synapses also occurred [16, 17]. Recent studies have demonstrated regeneration of ribbon synapses between afferent dendrites and inner hair cells [18, 19]. Actually, in our previous study to examine roles of prostaglandin E-type receptor 4 in mice, noise exposure after topical application of an antagonist prostaglandin E-type receptor 4 caused approximately 20 % loss of outer hair cells, but spiral ganglions and inner hair cells were well maintained [20]. Although of limited damage to outer hair cells, severe hearing loss was found in ABR measurements. In such a case, synaptic dysfunction between inner hair cells and spiral ganglion neurons might be involved. More recently, degeneration of ribbon synapses in the inner hair cells has been shown an initial change in age-related cochlear damage [21].
Proteins associated with the functionality of ribbon synapses have been reported. Myosin VI, bassoon, vglut3, Cav1.3, synaptotagmin 4, and otoferlin have close relation with ribbon synapse function, especially exocytosis [2]. The lack of myosin VI in mutant mice resulted in a dramatic decrease in synaptic exocytosis of inner hair cells [22]. Mouse mutants for the presynaptic scaffolding protein bassoon, which anchors synaptic ribbons at the active zone of inner hair cells, showed moderate hearing loss [23]. Inner hair cells in vglut3-null mice lack glutamate release, despite unaffected synaptic vesicular fusion [24]. In addition, vglut3-null mice showed degeneration of spiral ganglion neurons. Mutations in vglut3 were detected in patients with auditory neuropathy [24]. Otoferlin has been proposed as a calcium ion sensor of synaptic exocytosis. In otoferlin-null mice, exocytosis of inner hair cells is defective [25]. Mutations in OTOF were also detected in patients with auditory neuropathy.
References
1.
Merchan-Perez A, Liberman MC. Ultrastructural differences among afferent synapses on cochlear hair cells: correlations with spontaneous discharge rate. J Comp Neurol. 1996;371(2):208–21.PubMedPubMedCrossRef
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


