Superantigens (SAgs) are derived from diverse sources, including bacteria, viruses, and human hepatic tissue. SAgs initially cause lymphocyte activation but then result in clonal deletion and anergy, leading to immune tolerance. They can also act as superallergens by stimulating a broad spectrum of mast cells and basophils in patients with allergic conditions. The newly described staphylococcal SAg-like proteins subvert innate immune function by several mechanisms, which are distinct from SAgs’ effects on lymphocytes and other acquired immune processes. There is mounting evidence to suggest that SAgs play a role in the pathophysiology of inflammatory airway disease. The pathophysiologic role of SAg-like proteins awaits clarification.
The concept of superantigens (SAgs) was first described in 1989 in reference to the polyclonal activation of T lymphocytes by the staphylococcal enterotoxin B. Since then, many other proteins with SAg properties have been reported. A new group of SAgs, which act as superallergens by stimulating mast cells and basophils with a wide range of immunoglobulin (Ig) E specificities, have been observed. In addition, a group of SAg-like proteins, which have multiple effects on the innate immune system, have been recently described.
It is now known that SAgs are derived from diverse sources, including gram-positive and gram-negative bacteria, viruses, and even human hepatic tissue. There is evidence that SAgs have actions not only on T lymphocytes but also more broadly within the immune system, affecting B lymphocytes, mast cells, basophils, and chemokine production. Of particular relevance to otorhinolaryngologists is the involvement of SAgs in the pathogenesis of inflammatory airway conditions, including allergic rhinitis, chronic rhinosinusitis with nasal polyposis (CRSwNP), and asthma.
This article aims to summarize the current knowledge of SAgs and their relevance to current clinical rhinologic practice and research.
Immune response to conventional antigens
To explain the actions of SAgs, the immune response to conventional antigens is very briefly outlined. Antigen-presenting cells (APCs), such as monocyte-macrophage cells, dendritic cells, or B lymphocytes, encounter an antigen, internalize and process it, and then present it on their cell membranes held within the antigen-presenting groove of major histocompatibility complex (MHC) class II proteins. Subsequent activation of a CD4 + helper T lymphocyte occurs when its cell surface receptor (T lymphocyte cell receptor [TCR]) interacts with the presented antigen–MHC II complex on the APC ( Fig. 1 ). This interaction, which requires CD4 as a coreceptor, is highly specific; only a few T-lymphocyte clones will possess the particular TCR capable of binding to a given antigen. The conventional antigen–MHC II interaction involves the variable regions of both the TCR’s α and β chains (TCRV α and TCRV β ). The result is a T-lymphocyte clonal response in which there is a selective activation and proliferation of less than 0.1% of the naive T-lymphocyte repertoire. Subsequently, some clones are deleted but others persist as memory T lymphocytes, which facilitate a more rapid and vigorous secondary immune response after reexposure to the same antigen.
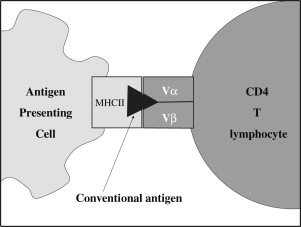
The humoral immune system also plays a role in the response to conventional antigens. Natural antibodies (usually IgM) act as a first line of defense by binding to antigens, which enter the circulation. In a manner analogous to the TCR-antigen interaction, B lymphocytes have membrane-bound immunoglobulins, which act as antigen receptors (B lymphocyte cell receptors [BCRs]). Antigen binds to a specific BCR, leading to a B-lymphocyte clonal response. B lymphocytes also present the processed antigen to T lymphocytes, and this interaction stimulates transformation of B lymphocytes into plasma cells and the synthesis of immunoglobulins specific for that antigen.
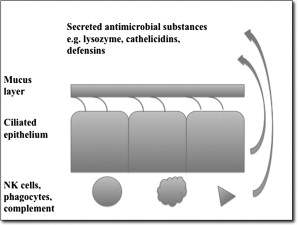
The innate immune system plays an important role in sinonasal mucosal defense and has several components ( Fig. 2 ).
The actions of SAgs on the immune system
The initial report of the effect of a staphylococcal SAg on T lymphocytes described a non–antigen specific activation. Subsequent research has shown that the biologic effects of these proteins are much more diverse. Although many of the effects of SAgs are proinflammatory, it is now understood that during the early stages of bacterial colonization or infection SAgs may in fact facilitate the avoidance or suppression of local immune mechanisms. In this way, SAgs may confer an evolutionary advantage on bacteria.
The classical SAgs are Staphylococcus aureus exotoxins. These proteins have 3 targets in the adaptive immune system: the TCR, the BCR, and the MHC class II proteins on APCs. In this manner, SAgs may be classified as T- or B-lymphocyte SAgs.
The Actions of SAgs on T Lymphocytes
SAgs are the most powerful of all known T-lymphocyte mitogens. Unlike conventional antigens, T-lymphocyte SAgs do not undergo processing by APCs. Instead, they possess structural features that allow them to bind directly and sequentially to the MHC class II proteins on APCs and then to the TCR’s β chain at a region that lies outside the conventional antigen–specific binding site ( Fig. 3 ). In this manner, the conventional antigen-specific T-lymphocyte activation is bypassed and a T-lymphocyte supraclonal response follows, involving more than 5% of the naive T-lymphocyte repertoire. This widespread activation may induce massive cytokine release from CD4 + lymphocytes (predominantly interleukin [IL] 2 and tumor necrosis factor α) and CD8 + lymphocytes (predominantly interferon gamma). In contrast to the response to conventional antigens, most of the activated T lymphocytes are subsequently deleted, sometimes to such an extent that their numbers decrease below original levels. This clonal deletion benefits both the host, by moving toward resolution of the inflammation, and the microbe, by allowing it to subvert the immune response, because the clones that are not deleted become anergic rather than becoming memory cells. In addition, SAgs activate CD4 + regulatory T lymphocytes, which then act to suppress the immune response via secretion of IL-10 and transforming growth factor β.
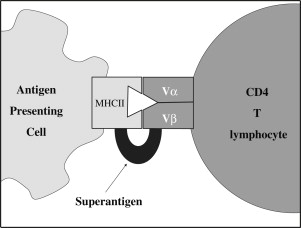
Different SAgs show different binding preferences for various MHC class II alleles and TCR profiles. However, the affinity of SAgs binding to MHC class II molecules is consistently more than 10 times greater than their affinity for binding to TCR. There exist low-affinity and high-affinity binding interfaces between SAgs and different MHC class II alleles. Of the human MHCs, HLA-DR tends to have the greatest affinity for SAg binding, followed by HLA-DQ, and then HLA-DP. With regard to TCR interactions, SAgs have been divided into 3 groups based on their specificity of binding to regions of TCRV β proteins. Human T lymphocytes have about 50 possible TCRV β regions, and each SAg may stimulate up to 20% of these regions.
Unlike conventional antigens, SAgs do not need CD4 as a coreceptor in the activation of T lymphocytes, and their structure does not change significantly after binding.
The Actions of SAgs on B Lymphocytes
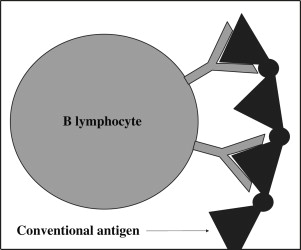
B lymphocytes have cell surface immunoglobulins, which interact with antigens without requiring presentation of the antigen by MHC class II proteins of APCs. B-lymphocyte SAgs bypass the conventional antigen–binding site of human immunoglobulin molecules. Conventional antigens bind to the hypervariable region of surface immunoglobulins ( Fig 4 ), whereas SAgs bind to certain areas of the variable regions of the heavy or light chain of membrane-associated immunoglobulin (which is the B lymphocyte equivalent of the TCR, Fig 5 ). These areas have been conserved over the course of human evolution so that a large proportion of B lymphocytes possess them. B-lymphocyte SAgs then activate BCRs by cross-linking them. The cross-linking is possible because the SAgs are composed of oligovalent or multivalent arrays of antigen-binding fragment (Fab)–binding domains. The result is a supraclonal B-lymphocyte proliferation, which is followed by cell death in some clones. In vivo studies have shown that administration of B-lymphocyte SAgs can deplete subsets of B cells, which have innatelike immune functions (such as secretion of natural antibodies). In particular, B1 and marginal zone B lymphocytes, which have innate immune functions, are inherently more sensitive to BCR-mediated signaling than other subsets of B lymphocytes, and strong stimulation results in apoptosis of these cells. B-lymphocyte SAgs exploit this tendency and cause deletion of these B-lymphocyte clones. There has been a speculation that this then leads to a weakened–host immune response to the bacteria.
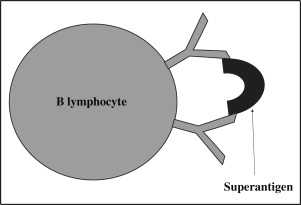
Different SAgs show different binding preferences for the areas within the variable region of the BCR. The prototypic B-lymphocyte SAg is staphylococcal protein A, which targets the Fab portion of the heavy chain of between 15% and 50% of polyclonal IgG, IgM, IgA, and IgE. In contrast, Peptostreptococcus magnus protein L (PpL) targets immunoglobulin light chains. S aureus enterotoxins A (SEA) and D are also B-lymphocyte SAgs, and they enhance the survival of the B cells to which they bind in vitro.
The Actions of SAgs on Chemokine Production
Chemokines are a superfamily of proteins produced by immune cells, which regulate innate and adaptive immune responses and are especially involved with leukocyte chemotaxis, including leukocyte adhesion to vascular endothelium, extravasation, and migration through tissue. Chemokines interact with cell surface receptors and one group, the cluster chemokines, binds to many different receptors.
There is evidence that staphylococcal SAgs are able to increase chemokine output in atopic dermatitis. SAgs may directly increase production of chemokine (C-C motif) ligand 1 (CCL1) and CCL18 by dermal dendritic cells, after these cells have interacted with T lymphocytes. CCL1, which binds to the receptor CCR8, may then promote survival of T lymphocytes and dendritic cells at the sites of inflammation. Also, SAgs may induce memory T lymphocytes to secrete cytokines, especially IL-31 that causes increased chemokine expression by keratinocytes. IL-31 is significantly associated with pruritis, which leads to scratching, skin trauma, and subsequent amplification of the inflammatory cascade.
The actions of SAgs on the immune system
The initial report of the effect of a staphylococcal SAg on T lymphocytes described a non–antigen specific activation. Subsequent research has shown that the biologic effects of these proteins are much more diverse. Although many of the effects of SAgs are proinflammatory, it is now understood that during the early stages of bacterial colonization or infection SAgs may in fact facilitate the avoidance or suppression of local immune mechanisms. In this way, SAgs may confer an evolutionary advantage on bacteria.
The classical SAgs are Staphylococcus aureus exotoxins. These proteins have 3 targets in the adaptive immune system: the TCR, the BCR, and the MHC class II proteins on APCs. In this manner, SAgs may be classified as T- or B-lymphocyte SAgs.
The Actions of SAgs on T Lymphocytes
SAgs are the most powerful of all known T-lymphocyte mitogens. Unlike conventional antigens, T-lymphocyte SAgs do not undergo processing by APCs. Instead, they possess structural features that allow them to bind directly and sequentially to the MHC class II proteins on APCs and then to the TCR’s β chain at a region that lies outside the conventional antigen–specific binding site ( Fig. 3 ). In this manner, the conventional antigen-specific T-lymphocyte activation is bypassed and a T-lymphocyte supraclonal response follows, involving more than 5% of the naive T-lymphocyte repertoire. This widespread activation may induce massive cytokine release from CD4 + lymphocytes (predominantly interleukin [IL] 2 and tumor necrosis factor α) and CD8 + lymphocytes (predominantly interferon gamma). In contrast to the response to conventional antigens, most of the activated T lymphocytes are subsequently deleted, sometimes to such an extent that their numbers decrease below original levels. This clonal deletion benefits both the host, by moving toward resolution of the inflammation, and the microbe, by allowing it to subvert the immune response, because the clones that are not deleted become anergic rather than becoming memory cells. In addition, SAgs activate CD4 + regulatory T lymphocytes, which then act to suppress the immune response via secretion of IL-10 and transforming growth factor β.
Different SAgs show different binding preferences for various MHC class II alleles and TCR profiles. However, the affinity of SAgs binding to MHC class II molecules is consistently more than 10 times greater than their affinity for binding to TCR. There exist low-affinity and high-affinity binding interfaces between SAgs and different MHC class II alleles. Of the human MHCs, HLA-DR tends to have the greatest affinity for SAg binding, followed by HLA-DQ, and then HLA-DP. With regard to TCR interactions, SAgs have been divided into 3 groups based on their specificity of binding to regions of TCRV β proteins. Human T lymphocytes have about 50 possible TCRV β regions, and each SAg may stimulate up to 20% of these regions.
Unlike conventional antigens, SAgs do not need CD4 as a coreceptor in the activation of T lymphocytes, and their structure does not change significantly after binding.
The Actions of SAgs on B Lymphocytes
B lymphocytes have cell surface immunoglobulins, which interact with antigens without requiring presentation of the antigen by MHC class II proteins of APCs. B-lymphocyte SAgs bypass the conventional antigen–binding site of human immunoglobulin molecules. Conventional antigens bind to the hypervariable region of surface immunoglobulins ( Fig 4 ), whereas SAgs bind to certain areas of the variable regions of the heavy or light chain of membrane-associated immunoglobulin (which is the B lymphocyte equivalent of the TCR, Fig 5 ). These areas have been conserved over the course of human evolution so that a large proportion of B lymphocytes possess them. B-lymphocyte SAgs then activate BCRs by cross-linking them. The cross-linking is possible because the SAgs are composed of oligovalent or multivalent arrays of antigen-binding fragment (Fab)–binding domains. The result is a supraclonal B-lymphocyte proliferation, which is followed by cell death in some clones. In vivo studies have shown that administration of B-lymphocyte SAgs can deplete subsets of B cells, which have innatelike immune functions (such as secretion of natural antibodies). In particular, B1 and marginal zone B lymphocytes, which have innate immune functions, are inherently more sensitive to BCR-mediated signaling than other subsets of B lymphocytes, and strong stimulation results in apoptosis of these cells. B-lymphocyte SAgs exploit this tendency and cause deletion of these B-lymphocyte clones. There has been a speculation that this then leads to a weakened–host immune response to the bacteria.
Different SAgs show different binding preferences for the areas within the variable region of the BCR. The prototypic B-lymphocyte SAg is staphylococcal protein A, which targets the Fab portion of the heavy chain of between 15% and 50% of polyclonal IgG, IgM, IgA, and IgE. In contrast, Peptostreptococcus magnus protein L (PpL) targets immunoglobulin light chains. S aureus enterotoxins A (SEA) and D are also B-lymphocyte SAgs, and they enhance the survival of the B cells to which they bind in vitro.
The Actions of SAgs on Chemokine Production
Chemokines are a superfamily of proteins produced by immune cells, which regulate innate and adaptive immune responses and are especially involved with leukocyte chemotaxis, including leukocyte adhesion to vascular endothelium, extravasation, and migration through tissue. Chemokines interact with cell surface receptors and one group, the cluster chemokines, binds to many different receptors.
There is evidence that staphylococcal SAgs are able to increase chemokine output in atopic dermatitis. SAgs may directly increase production of chemokine (C-C motif) ligand 1 (CCL1) and CCL18 by dermal dendritic cells, after these cells have interacted with T lymphocytes. CCL1, which binds to the receptor CCR8, may then promote survival of T lymphocytes and dendritic cells at the sites of inflammation. Also, SAgs may induce memory T lymphocytes to secrete cytokines, especially IL-31 that causes increased chemokine expression by keratinocytes. IL-31 is significantly associated with pruritis, which leads to scratching, skin trauma, and subsequent amplification of the inflammatory cascade.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


