Fig. 35.1
The facial profile of a 3-month-old boy (a) and his father (b). Proportional differences between the facial and brain skull of father and son. The baby face shows smaller vertical dimensions, a less frontal projection of the nose and a larger nasolabial angle
In the newborn, the cartilaginous and bony nasal skeleton demonstrates a few specific aspects. Septal cartilage and upper lateral cartilages on both sides form the three-dimensional septodorsal cartilage, a supporting structure for the nasal dorsum resembling a T-bar configuration (Poublon et al. 1990). The cartilaginous septum is based on the sphenoid (Fig. 35.2 ). The upper lateral cartilages extend under the nasal bones to merge with the cartilaginous anlage of the anterior cranial base. The nasal bones are fibrously connected in the sutures to the frontal and maxillary bones. At their ventral rim, the periost of the nasal bones is firmly connected to the perichondrium of the underlying upper lateral cartilages. The nasal bones are product of extra-cartilaginous ossification of cephalic mesenchyme. The first anlage of the vomer is also represented by islands of mesenchymal bone formation reaching from palatal bone to cartilaginous septum with extensions of ossifying mesenchyme along both sides of the septum cartilage. Enchondral ossification of the septum cartilage may be observed as early as the first months after birth near the anterior cranial base (Van Loosen et al. 1988). The cartilaginous septum shows a specific pattern of thinner and thicker zones; the thickness of the septum was found to vary between 0.4 and 3.5 mm. Two areas of thick cartilage are extending from the sphenoid in anterosuperior direction to the nasal dorsum (sphenodorsal zone) and the anterior nasal spine (sphenospinal zone), respectively (Fig. 35.2c). The thinnest part is found anteriorly between these two zones and the slightly thickened caudal rim.
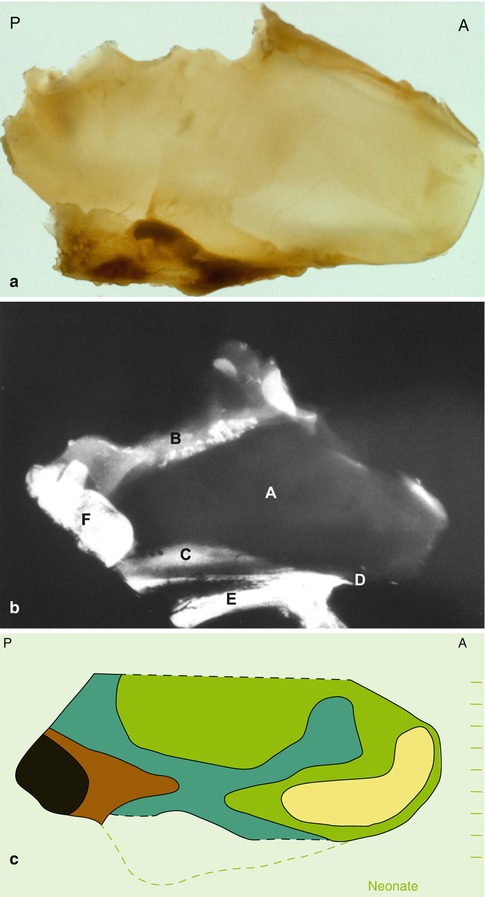
Fig. 35.2
Anatomical specimen of neonatal cartilaginous septum: (a) indentations at the border with the anterior cranial base concur with ossified parts lost during preparation; (A) anterior, (P) posterior. (b) Lateral radiograph of an anatomical specimen of a human neonate including the nasal skeleton, adjacent skull base and upper jaw. (A) Cartilaginous nasal septum, (B) lamina cribrosa, (C) vomeral wing, (D) anterior nasal spine, (E) palate, (F) sphenoid. (c) Schematic representation of the thinner and thicker zones in the human septum (superior part of the septum, adjacent to the anterior cranial base not included). The thinnest part (yellow, 400–500μ) is bordered by the columellar rim (light green, 500–1,500μ); sphenodorsal and sphenospinal zones of thicker cartilage (blue-brown, 1,500–3,000μ); sphenoid (black)
In young children the cartilaginous nasal septum is based on the sphenoid, whereas the upper lateral cartilages extend under the nasal dorsum to merge with the cartilaginous cranial base. Dimensions and anatomical features change with increasing age.
In young rabbits a similar pattern of thicker and thinner areas has been demonstrated in the elongated septum (Tonneyck-Müller and van der Werf 1982, 1984). The thicker areas play a specific and important role in the postnatal development of nose and upper jaw as will be discussed in the paragraph on acquired malformations of the septum. Also in young rabbits, the septal and upper lateral cartilages form a T-bar based on the anterior skull base.
35.1.3 Midfacial Development from Neonate to Adolescent
In children the development of the cartilaginous nasal skeleton is a complex process including proliferation of chondroblasts, increase of intercellular matrix, tissue maturation and partial regression and enchondral ossification of the septodorsal cartilage. The dimensional growth of the septum cartilage shows its highest rate in the newborn and is slowing down gradually after the age of 2 years (van Loosen et al. 1996). From that time onwards, formation of new cartilage continues but is balanced by simultaneous loss of cartilage through enchondral ossification (Verwoerd and Verwoerd-Verhoef 2007). Mitotic activity of chondroblasts and expansion of the intercellular matrix might compensate for the loss of cartilage by ossification till the ratio between bony and cartilaginous parts has been changed to its definite state (Fig. 35.3). Consequently, the sagittal dimensions of the bony perpendicular plate are increasing relative to the cartilaginous part of the septum (Schultz-Coulon and Eckermeier 1976). The growing perpendicular plate will intervene between septal cartilage and sphenoid. The septum cartilage is later firmly connected to the thickened caudal rim of the perpendicular plate. The junction of cartilaginous septum and perpendicular plate – an important surgical landmark – will change from an intracranial position in young children to the extracranial location at the anterior margin of the nasal bones in adults. The basis of the septum cartilage shifts from the sphenoid to the anterior rim of the perpendicular plate (Fig. 35.4a).
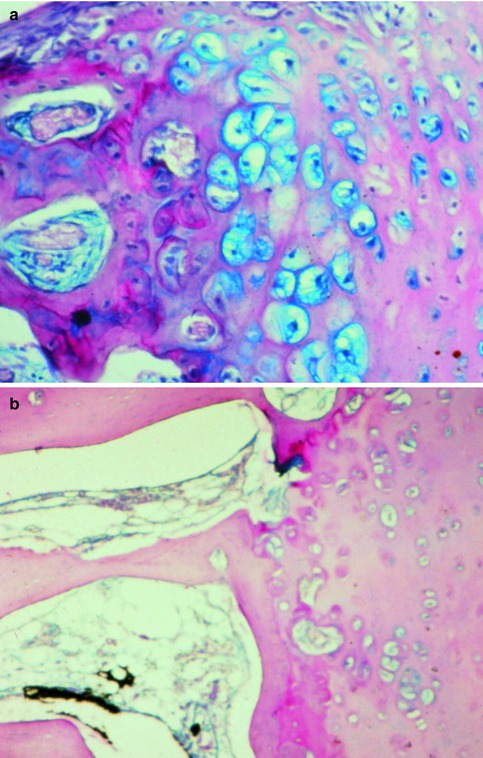
Fig. 35.3
The interface between septal cartilage (right) and bony perpendicular plate (left); (a) active enchondral ossification in a young child; (b) no signs of ossification in the adult stage
The vomeral bone, which first anlage was present in the neonate will develop into a definite part of the osseous nasal skeleton (Verwoerd et al. 1989a). The vomer is enclosing the basal rim of the septum cartilage by two bony layers (vomeral wings) which converge inferiorly in the medial, unpaired bony plate separating the inferior part of the nasal cavities and extending in posterior direction to the choanal edge of the nasal septum. The moment at which the ossifying front of the perpendicular plate should reach the vomeral wings is not clearly defined and may even surpass the age of 10 years, while an overlap between wings and plate has been found in adolescence. Cartilage may remain present in the bony canal formed by vomeral wings and perpendicular plate (vomeral tunnel) for a longer period and extend to the sphenoid, as sphenoid tail, but ultimately ossify in most individuals. Asymmetry of the vomeral wings (the ala on one side larger than on the other) may be observed when the sphenoid tail bulges out into one nasal cavity, sometimes in combination with a vomeral spine (Fig. 35.4b). Variations in the septovomeral junction are very common and a symmetrical development is exception rather than rule. In a study on human fetuses of 5 months old, it was observed by Takahashi that around 25 % demonstrated an abnormality of the septovomeral junction which could increase to almost 40 % at birth (Takahashi 1987). These deformities were ascribed to ‘an imbalance of the “overdeveloping” septum (cartilage) and the pressure of the surrounding structures’, the last mentioned being the developing bony facial skeleton.
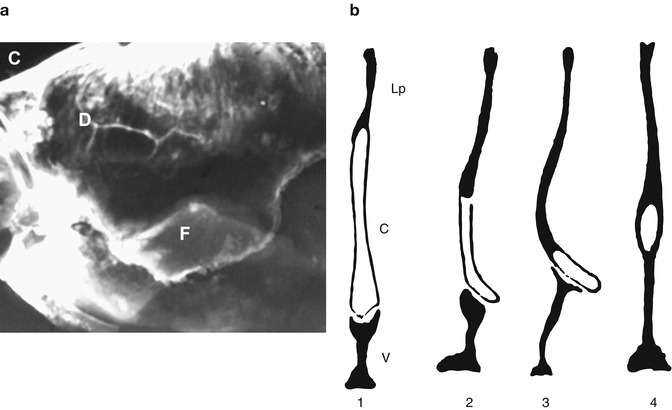
Fig. 35.4
Detail of a human nasal septum (17 years of age); (a) a lateral radiograph, with the overlap of vomeral wing and perpendicular plate; [C] anterior skull base, [D] lamina perpendicularis, [F] vomeral wing; (b) schematic representation (in frontal sections) of various modalities of the septoethmoidovomeral junction (after Takahashi 1987); [1] normal situation (Lp = lamina perpendicularis, c = cartilage, v = vomer), [2] asymmetrical development of vomer and cartilage, [3] asymmetrical development of vomer with formation of a vomeral spine, [4] sphenoid tail
Progressive ossification of the septum cartilage results in an expanding perpendicular plate starting from the area of the anterior cranial base into ventrocaudal direction. The ventral rim of the perpendicular plate shifts gradually inferiorly and, therefore, is in children not a reliable point of orientation in relation to the anterior skull base. The junction between cartilagenous septum, perpendicular plate and vomer may demonstrate considerable variation but is thought to have been established between 10 and 14 years of age.
The length of the upper lateral part of the dorsoseptal cartilage will be reduced on both sides to finally around 5–8 mm. The progress of this reduction shows individual variation. While the dimensions of the nasal skeleton increase in size, the relation between cartilaginous and bony parts is going to be altered. This remodelling will result in a more anterior position of the cartilaginous part of the septum. It is an important issue which should be understood by the doctor at diagnosis and before treatment of the child might be started.
Anatomical data do not give information pertinent to the developmental mechanisms of the facial skull. Animal experiments are necessary to analyse these morphogenetic mechanisms and the way they are affected by injury or surgery.
35.1.4 Postnatal Development of the Midfacial Skeleton in Mammals
The skulls of mammals demonstrate essentially similar components (Fig. 35.5). Various components however may show very different dimensions in different species. An example is the proportion between the skeletal components of the upper jaw. In the human skull, the premaxilla is small compared with the maxilla, whereas in rodents, like the rabbit, the anteroposterior dimensions of the premaxilla exceed those of the maxilla. In children, the osteogenic activity of the premaxillo-maxillary suture is restricted to the first years of life contrary to rabbits in which sutural growth contributes to lengthening of the upper jaw and continues till sexual maturity.
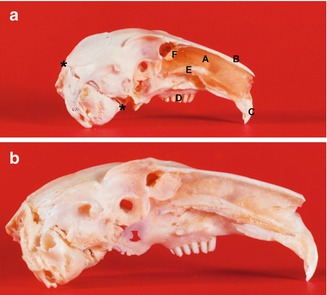
Fig. 35.5
Lateral aspect of the rabbit septum and skull (after removal of the right part of the nose and upper jaw) at the age of 4 (a) and 24 weeks (b) demonstrating the ‘extra’ growth of the nasal skeleton and upper jaw up to the adult stage, compared to the dimensional growth of the brain skull. *-* Line between lambdoid suture and spheno-occipital suture, (A) cartilaginous nasal septum, (B) nasal bone, (C) incisors, (D) molar complex, (E) vomeral wing, (F) perpendicular plate
Also in young rabbits the septodorsal cartilage extends under the nasal bones to the anterior cranial base, later followed by a reduction of the posterior part of the cartilage leaving only the most anterior part in situ. The cartilaginous septum remains in direct contact with the sphenoid. Only a small part will demonstrate ossification to form the perpendicular plate. Obvious differences compared to the human anatomy are demonstrated by the vomer. The vomer is a product of mesenchymal ossification along both sides of the cartilaginous septum and between the palatal bone and the inferior margin of the septum. The cartilaginous septum, anteriorly connected to the anterior nasal spine, shows thinner and thicker parts similar to those described for the human nasal septum (Fig. 35.6). A centro-anterior area of thin cartilage is surrounded by sphenodorsal and sphenospinal zones of thick cartilage and anteriorly bordered by a slightly thickened anterior rim. In growing rabbits, it was demonstrated that the ‘extra’ growth of nose and maxilla depends primarily on growth of the septodorsal cartilage (Fig. 35.7). The sphenodorsal zone of thicker cartilage is responsible for lengthening of the upper part of the T-bar, the upper lateral cartilages and indirectly of the overlying nasal bones. These upper laterals were found to stabilise the growing septum cartilage in a median position. Also in the rabbit the extension of the lateral cartilages under the nasal bones will be gradually reduced by ongoing regression (Fig. 35.8). Enlargement of the sphenospinal zone results in an increase in length and thus gradual shifting of the lower part of the septum and upper jaw. In the rabbit, the perpendicular plate will be limited to the most posterior part of the septum, whereas the majority of the septum remains cartilaginous. Also in the mouse, the cartilaginous septum increases in length much more rapidly than could be explained by caudal growth, implying that interstitial expansion is the more important contributor to septum development (Wealthall and Herring 2006).
Equally important as the septodorsal cartilage is for the growing midface are the sutures and their bone formation for the facial skeleton, the nose included.
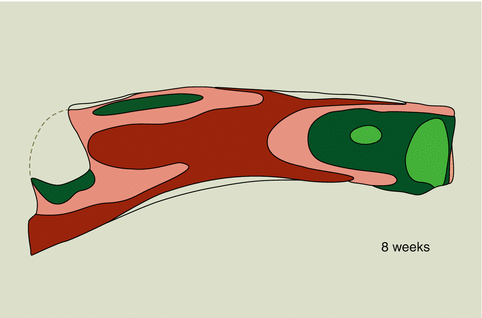

Fig. 35.6
Schematic presentation of regional differences in thickness of the cartilaginous nasal septum in an 8-week-old rabbit. The sphenospinal zone of thick cartilage extends from the sphenoid to the anterior nasal spine, whereas the sphenodorsal zone is extending under the nasal dorsum. In rabbits, the lower lateral cartilages have a common medial crus, which is connected by a thin cartilaginous membrane with the slightly thickened anterior rim of the ‘real’ cartilaginous septum; this thickened rim and the sphenodorsal and sphenospinal zones of thick cartilage enclose an area of thin cartilage; l.green: 50–250μ, d.green: 250–450μ, l.red: 450–650μ, d.red: 650–850μ
Fig. 35.7
Graphic representation of the ‘extra’ growth of the midfacial skeleton (with grid code) of the rabbit skulls between 4 and 24 weeks after birth; the line connecting lambdoid suture and the spheno-occipital suture (viz. Fig. 35.5a) has been made equal for both series; elongation of the upper jaw and the nose and forward shift of the molar complex result in the adult proportions between facial and brain skull
Fig. 35.8
Schematic representation of the regression of the upper lateral cartilage from (a) 4 to (b) 24 weeks; the left nasal bone is removed (Courtesy of Dr R.M.L. Poublon)
35.1.5 Dimensional Growth of the Nose and Maturation
Postmortem anatomical studies suggested a phase of rapid growth directly after birth with a gradual deceleration after 5 years with the greatest velocity in the first 2 years (van Loosen et al. 1996, 1997). Conclusions drawn from a study in the Aegean Region of Turkey revealed that nasal height and nasal bridge length reached full maturation in females already at 12 years of age and in boys around 15 years (Akgüner et al. 1998). Nasal growth has further been studied by measuring cohorts of children and calculating ‘standards’ for various age groups, differentiating for boys and girls. Next to these horizontal studies, a few vertical studies have been published based on measurements in the same child at increasing ages. Such a vertical study demonstrated in boys a period of accelerated growth, most frequently observed around the age of 13 years (Meng et al. 1988). In young girls, periods of accelerated growth were found to occur between the age of 6 and 8 years. Growth spurts have not been demonstrated in horizontal studies including large cohorts of children. In the last two decennia, most data on postnatal growth of the maxilla and the nose have been derived from lateral cephalographs in children of 7 years and older (Meng et al. 1988; Farkas et al. 1992; Zankl et al. 2002; Ochoa and Nanda 2004). In a recent review, the steepest descending slope of midfacial growth velocity is reported to take place at the average age of 13.4 years for adolescent girls and 14.7 years for boys (van der Heijden et al. 2008). It was suggested that 98 % of white, adolescent girls are ‘nasally mature’ at the age of 15.8 years and 98 % of the boys at the age of 16.9 years.
35.2 Pathophysiology of the Growing Nasal Skeleton
35.2.1 Congenital Anomalies
35.2.1.1 Midfacial Clefts and the Nasal Septum
Craniofacial growth is a dynamic process balancing form and function. This equilibrium is easily disturbed at various levels by passive or active change of one of the mechanisms involved, prenatally or postnatally. The impact of the interaction with the adhering muscles on the developing midfacial skeleton has been recognised in recent years (Stricker et al. 1990). A spectrum of midfacial malformations may be observed as part of a syndrome or as solitary deformity which may point to an interaction between growth of the nasal septum and the premaxilla-maxilla. One of the most well-known congenital facial deformities is the cleft lip, alveolus and palate. In adult human skulls with facial clefts, which were known to be untreated, a specific pattern of growth disturbances could be observed (van Limborgh 1964; Atherton 1967). Unilateral clefts showed a deviation of the premaxilla to the non-cleft side, whereas the maxillary part on the cleft side had collapsed medially and fell behind compared to the non-cleft side (retrognathism). In addition, these skulls showed a specific malformation of the nasal septum with (a) a deviation of the perpendicular plate to the cleft side and (b) a disjunction between perpendicular plate and vomer (Fig. 35.9). The vomer, only connected to the palatal margin of the cleft, tends to a more horizontal position to meet the deviated perpendicular plate, suggesting a broadening of the nasal floor on the non-cleft side. From this study, it was concluded that these growth anomalies of the maxilla and nasal framework were part of a cleft syndrome, specific for each type of cleft (Verwoerd et al. 1989a, b). Collapse of the upper alveolar arch and maxillary retrognathism were also observed in adult members of a North-Borneo tribe who were born with cleft lip, alveolus and palate and did not undergo surgical treatment (Innis 1962). In a study on lateral cephalograms of untreated unilateral cleft patients with superimposed images of cleft and non-cleft side, the abnormal position of the maxilla could not confirmed, most probably due to the overprojection of the non-affected side (Capelozza et al. 1993).
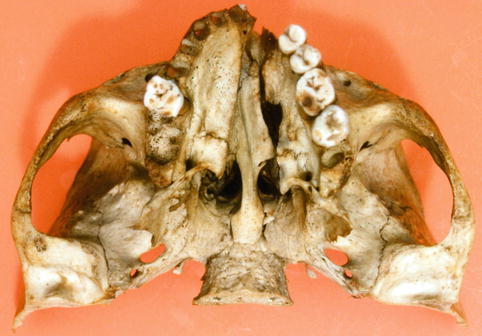
Fig. 35.9
Caudal view of a human skull with unilateral cleft of alveolus and palate (Dept. of Anatomy and Embryology, Amsterdam UMC). The maxillary segment on the cleft side demonstrates a medial collapse and a retroposition (asymmetrical position of the left and right tuber maxillae); deviation of vomer and premaxilla to the non-cleft side
Whether a combination of midfacial anomalies associated with a cleft alveolus and palate might represent a syndrome of craniofacial anomalies developing as reaction to a cleft was investigated in growing rabbits (Verwoerd et al. 1979a). Unilateral clefts of alveolus and palate, produced surgically in young growing rabbits, appeared to affect the further development of nose and (pre)maxilla into adulthood (Fig. 35.10a). On the cleft side the molar complex, lacking a connection with the growing septum, does not move forward and will show – in the adult stage – a retroposition (retrognathism) compared to the non-cleft side. Here, the premaxilla, maxilla and nasal bones will grow to normal length and gradually rotate and deviate to the non-cleft side as previously reported for human skulls (van Limborgh 1964). The results of these animal experiments indicate that the cleft syndrome as identified in ‘untreated’ human skulls with facial clefts should be considered an adaptation to the altered developmental mechanics in the developing midfacial skeleton.
The interaction between the growing cartilaginous and bony skeleton is modified when midfacial sutures are missing as in the presence of facial clefts; similar cleft syndromes were found in animals and untreated patients.
Another interesting observation has been done when in young rabbits the premaxillary-maxillary suture was resected and replaced by non-sutural bone (Fig. 35.10b). Following this intervention normal lengthening of the ipsilateral upper jaw failed. Secondary effects were a progressive deviation of the premaxilla to the nongrowing side and an excessive forward shift of the molar complex on the operated side. These observations confirm disappointing midfacial development as observed after osteoplasty of an alveolar cleft in young children. The outcome of the above-mentioned experiments refers to the role of the cartilaginous nasal septum in postnatal development of the upper jaw. The growing and lengthening of the cartilaginous nasal septum, connected on both sides to the premaxilla-maxilla, seems to be responsible for lengthening of the upper jaw and, secondly, for a shift in anterior direction of the upper jaw relative to the cranial base. In the presence of a unilateral cleft, the ‘mechanical’ balance between both sides is disturbed.
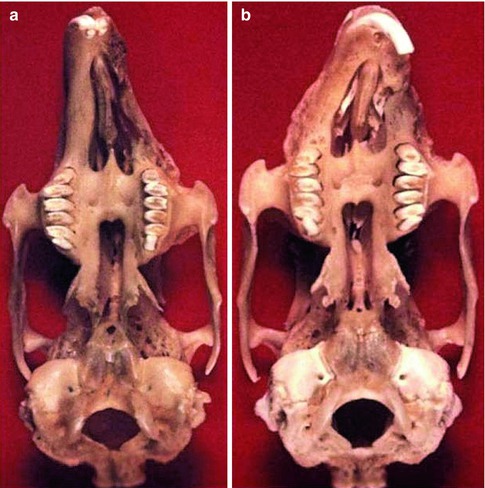
Fig. 35.10
Caudal aspect of the skulls of adult rabbits (24 weeks of age) with maldevelopment following surgical procedures at the age of 4 weeks: (a) following a surgical production of unilateral cleft lip and alveolus (resection of the premaxilla-maxillary suture): deviation of the premaxilla from the midline, retroposition of the maxilla, zygoma and pterygoid process on the cleft side; (b) after a similar cleft has been made and closed by primary osteoplasty: marked deviation to the operated side with shortening of the upper jaw, anteroposition of the maxilla including molar complex, zygoma and pterygoid process
Although the results of orthodontic and surgical treatment of facial clefts are improving, the secondary cleft nose in these patients can demonstrate some of the following features: asymmetry of tip, columella, nostril, ala and nostril floor; deflection of the caudal part of the septum cartilage to the non-cleft side; stenosis of the vestibule on the cleft side; hypoplasia of the maxilla on the cleft side; variable collapse of the maxilla with asymmetry of the piriform aperture ; underdevelopment of the maxilla with retroposition of the anterior nasal spine; deviation of the cartilaginous and bony nasal dorsum; and deviation of the posterior part of the nasal septum to the cleft side (Verwoerd et al. 1995).
35.2.1.2 Congenital Malformation of the Nose
Nasal congenital anomalies are extremely rare and range from bifidity of the nasal tip or dorsum to nasal aplasia with or without proboscis (Stricker et al. 1990). Reports on nasal dysplasia, which is mostly indicating a unilateral malformation, are scattered through the literature. The nasal cavity is missing and pneumatisation of the maxillary, ethmoidal and frontal sinuses has failed. Exploration reveals no cartilage, but just solid bone. The affected side of the maxilla is hypoplastic and regularly associated with other malformations such as cleft lip/palate or coloboma. The last variety to be mentioned is a duplication of the nasal dorsum, which can occur in different forms and as part of a syndrome (Fig. 35.11).
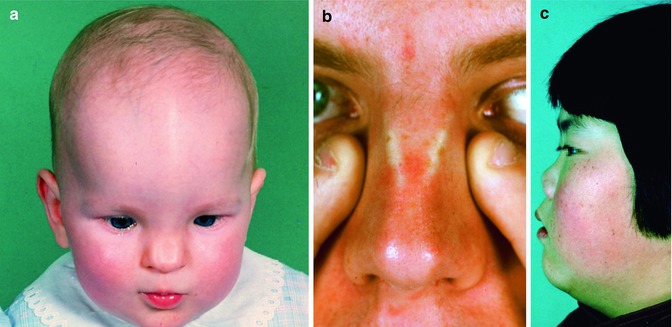
Fig. 35.11
Congenital anomalies of the nose. (a) Incomplete fusion of the left and right anlage of the cartilaginous and bony nasal skeleton in a 3-year-old girl and (b) an adult man. (c) Underdevelopment of the cartilaginous and bony nasal pyramid
35.2.2 Acquired Anomalies of the Nose
35.2.2.1 The Nose of the Neonate
Various studies refer to the importance of the nasal septum for midfacial development. In humans, intrauterine exposure to warfarin appeared to cause early calcification of the septal cartilage; subsequent nasal and midfacial hypoplasia was demonstrated in a cephalometric study, suggesting that even in utero midfacial growth is retarded by interference with the nasal septum (Howe et al. 2004).
The neonate’s nose may show a slight or more pronounced deviation, with a luxation of the lower septal border into the nasal cavity, acquired during the passage through the birth canal (Fig. 35.12). In most cases this anomaly will restore spontaneously but infrequently the deformity has to be corrected by manipulation of the neonatal nasal septum which is still mostly cartilaginous from sphenoid to columella. Already in 1978, Gray found some type of septum anomaly in 57 % of 2,380 neonates (Gray 1978). Septal deformity was explained by Takahashi to be an inevitable condition resulting from the autonomic growth force of the septum cartilage (Takahashi 1988).
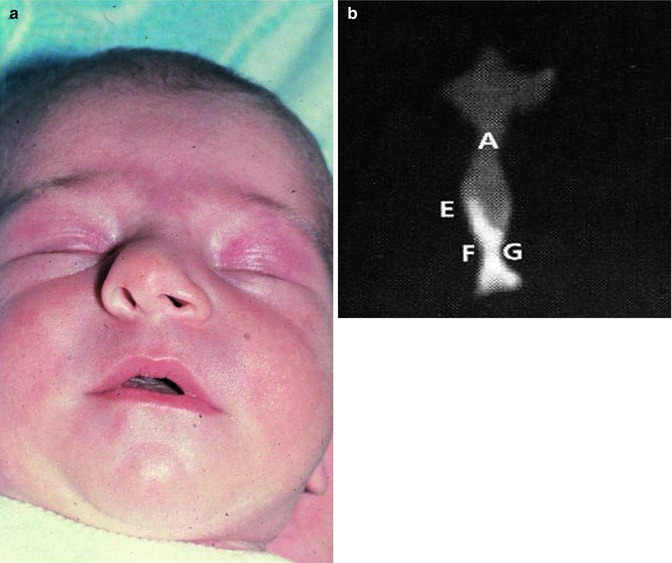
Fig. 35.12
A newborn baby with a deviation of the cartilaginous nasal dorsum and columella (a) due to a bending of the cartilaginous septum at the weak zone superior to the thick basal rim or a dislocation of the basal rim from the vomeral gutter, acquired during the passage through the birth canal. (b) Asymmetrical anlage of the vomer in a CT scan (frontal plane) of a neonatal septum. (A) Cartilaginous nasal septum, (E) ala vomeris, (F,G) inferior parts of the vomer
35.2.2.2 Trauma at a Young Age; Bone and Cartilage Lesions
Any kind of mechanical or surgical trauma at a young age, destroying the septodorsal cartilage, initiates an irreversible disturbance of the cartilage from its genetically based developmental orientation. A lesion may vary from fracture and dislocation, with later deviation, to haematoma and abscess formation and loss of septum cartilage, with various associated sequelae. The regional differences in thickness of the septum cartilage, described above, seem to explain that fractures preferably occur at sites with the least mechanical strength and thus the thinnest parts of the septum. Septal deviations could, therefore, be related to the ‘weak’ areas (Mladina 2000). Dislocation of the caudal end of the septum, the most frequent disorder in young children, is caused by a fracture running through the thin area lying cephalic of the columellar rim. The thin area above the thick basal rim is another vulnerable fracture-prone region. This preference of specific fracture lines was described by several authors in children as well as adult patients; it was demonstrated in the so-called C-fracture running horizontally and superiorly along the thick basal rim of the septum, then more horizontally through the thin portion of the perpendicular plate, while ending in the thinner zone just under the nasal dorsum (Harrison 1979; van Velzen et al. 1997). Another weak point is the area of thin cartilage, semi-parallel to the caudal rim and just anterior to the connection between septum and upper lateral cartilages, which may easily fracture when the caudal end of the septum gets dislocated (viz. Fig. 35.2c).
35.2.2.3 Fractures: Direct Effects and Follow-Up
Although injuries of the nose appear to occur frequently in young children – and the incidence seems still to be growing – only a relative small number is seen by the general practitioner, and even less are diagnosed by an ORL specialist. Consequently the long-term follow-up of these cases is rare and certainly no routine. In the years following such a ‘minor’ trauma, however, a progressive deformity of the midface may be observed with increasing deviation or distortion of the nose, eventually in combination with an underdevelopment of the maxilla, while the final effects can only be defined after the adolescent growth spurt.
After a traumatic accident, fractured parts of the bony midface and the septum will lead to, often disproportionate, disfigurement of the facial profile. In particular, the less obvious lesions of the cartilaginous septum are seldom diagnosed and during further growth the fractured parts will bend and dislocate increasingly, thus leading to spine formations, local overlapping edges and finally to severe deviations (Manning 1999). A midfrontal trauma may cause multiple fractures with larger and smaller pieces of cartilage resulting in a highly distorted septum and finally an underdeveloped and distorted nose (Fig. 35.13). The direction of deviations is mostly unpredictable, because it depends on the direction of the deforming force and on the reaction of the traumatized tissues involved. Captivating evidence for the significance of the nasal septum in midfacial growth is supplied by a rather large study of 21 monozygotic twins ranging in age from 18 to 22 years by Grymer (Grymer et al. 1991). In only one of each twin pair, a deformity was found in the anterior part of the septum. The nose of the affected siblings was demonstrated by cephalometry to be shorter than in their ‘normal’ other half.
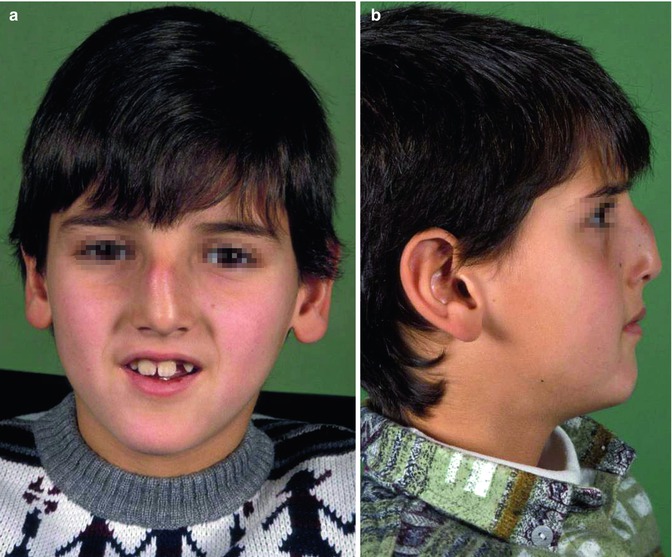
Fig. 35.13
Underdevelopment and deviation of the cartilaginous nasal pyramid, multiple and irregular deviations of the cartilaginous septum after a previous injury; (a) boy at the age of 8 years and (b) 13 years. Could a progressive anomaly have been prevented?
35.2.2.3.1 Animal Experiments
Poor wound healing of cartilage is the main reason for the occurrence of disorders of the nasal skeleton, with or without nasal airway obstruction. Experimental studies in young rodents and other mammals have consistently demonstrated that removal of full-thickness (cartilage + mucosal lining) portions of the growing nasal septum may result in growth retardation of nose, maxilla, premaxilla and palate (Kremenak and Searls 1971; Verwoerd et al. 1980; Sarnat 1983; Pinkston et al. 1995). Later studies with larger series of experiments in young rabbits using submucosal techniques could fill in some open ends of these earlier investigations as discussed in Sect. 35.2.2.4 (Verwoerd and Verwoerd-Verhoef 2011).
The growth of the rabbit skull was studied from 0 to 24 weeks after birth. The facial skeleton appeared to grow faster and over a longer period than the brain skull like in humans (Fig. 35.7). The growth rate increased rapidly in the first 8 weeks with a gradual decline thereafter till around the 22nd week of age. Twenty-four weeks was the final date of the experimental period. To achieve a complete image of the significance and developmental role of the bony and cartilaginous parts, the first experiments were performed on the nasal bones, maxilla and lateral nasal wall. Transverse osteotomy of the nasal bone anterior to the frontonasal suture on one or two sides did not disturb nasal growth in length nor did extirpation of the frontonasal suture alter the growth of the snout (Sarnat 2008). The snout showed no asymmetry, and a small bony defect remained only present in a small minority. Even subtotal resection of the nasal bone did not result in growth disorders as long as the underlying upper lateral cartilage was undamaged (Verwoerd et al. 1980). The same restriction applied to partial resection of the lateral nasal wall, which in rabbits can only be performed through an opening of the nasal cavity from above after mobilisation of the nasal bone without disturbing the upper lateral cartilage on that side (Verwoerd-Verhoef and Verwoerd 2003). Although the FESS procedure in children is not completely analogous, it was demonstrated that enlargement of the entrance to the maxillary sinus via this method did not disturb the normal growth of the rabbit’s nose.
Experiments including resection of the premaxillo-maxillary suture(s), also in combination with resection of the midpalatal suture – comparable to the cleft upper jaw and palate – gave similar results as the cleft syndrome described for the human patients and skulls (see Sect. 35.2.1.1). The retroposition of the maxillary portion on the cleft side is explained by the failing connection to the growing septum at the anterior nasal spine. Consequently, the septum will only pull forward the premaxillary complex on the unaffected side and gradually deviate to that side. Malocclusion is the result. Closure of alveolar defect with non-sutural bone, like it was performed in children with cleft alveolus and palate in the 1970s and 1980s, demonstrated clearly that the development of the midface became severely disturbed (Verwoerd et al. 1979a).
35.2.2.4 Septum Haematoma and Abscess: Loss of Septum Cartilage
From many human case studies described in the literature, it is evident that the sequelae of a nasal injury are frequently a septum haematoma and, due to contamination and infection, a subsequent septum abscess. A haematoma may mostly, when not treated adequately, and an abscess may invariably cause destruction of the nasal cartilage leading to septum perforation or a submucosal defect of the septum cartilage. Saddling deformity with loss of tip support and growth retardation with a short nasal dorsum are then regular findings (Fig. 35.14a, b). Grymer and Bosch reported about one twin sibling, who endured septum destruction due to a septum abscess at the age of 7 years and was followed up with its twin for more than 10 years (Grymer and Bosch 1997). Despite the treatment with homologous cartilage from the tissue bank, the affected boy developed a severe saddle nose with upward displacement of the anterior part of the maxilla, decreased nasal projection and a retrognathical position of the maxilla compared to the unaffected sibling. This observation suggested that the nasal septum has a great influence on midfacial development determining vertical as well as anteroposterior maxillary growth. And secondly, that bank cartilage grafting is not adequate in preventing malformations of the midface during growth. Patients with a history of facial trauma develop progressive nasal deformation and/or nasal obstruction but the younger the age of the child the more impact such a lesion may have in the long run (Pirsig 2000). Also the extent of the injury – fracture, haematoma, abscess and perforation, surgical treatment – will play a role in the final result after the adolescent growth spurt. And these results are often disappointing, thus many of these patients have to undergo a second or even more surgical procedures later in life (Derkay 1999).
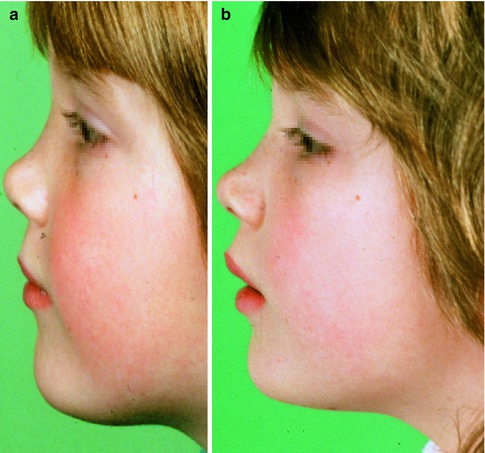
Fig. 35.14
Underdevelopment of the cartilaginous and bony nasal pyramid after a previous septal abscess; (a) girl at the age of 5 years and (b) 10 years. Increased nasolabial angle, with a prominent tip (supported by the alar cartilages)
From a series of 241 children with a history of nasal injury, 40 % was diagnosed with pathological septal findings, with a longest follow-up period after trauma of 10 years (Blahova 1985). Septum deviation associated with snoring and abnormal rhinomanometric values appeared in 55 % of the patients. In a group of 16 paediatric patients, ranging from 2 to 14 years, major sequelae (62,5 %) like dorsum, tip or pyramid deformation, septum deviation with nasal obstruction, functional vault deformity and septum perforation were observed, whereas the other patients (37,5 %) all showed minor sequelae without any airway compromise (Alvarez et al. 2000). Another consecutive series of 20 children (2 months–15 years) were admitted to the hospital for treatment of nasal trauma followed by nasal abscess (12 patients) universally associated with destruction of the septum cartilage (Canty and Berkowitz 1996). Nasal obstruction, however, was the most common symptom (19 patients). Recently a structured review of PubMed, EMBASE and the Cochrane Databases comprising 81 citations, dating from 1920 up to now, regarding nasal septal abscess was published (Alshaikh and Lo 2011). It was concluded that it is a serious condition that necessitates urgent surgical management in order to prevent severe complications, and even the growing child should be treated with early reconstruction essential for normal development of the midface. In this whole sequence, the lack of knowledge of the developmental mechanics is part of the problem.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


