Neonatal Ophthalmology: Ocular Development in Childhood
Kammi B. Gunton
THE NEONATAL EYE is one of the most fully developed sensory organs. Despite its similarity to the adult eye, there are tremendous changes that will occur in the refractive correction, axial length, shape of cornea, color of iris, pupillary responses, and retinal and neurologic development during childhood. Understanding this process is vital to the appropriate care of children’s eyes. This chapter focuses on the anatomic changes in the eye and orbit during infancy and through adolescence. Intraocularly, the anterior segment, retina, and optic nerve undergo rapid changes within the first year of life to allow the development of clear images on the retina. Neurologic development then allows the subsequent processing of the retinal image. Externally, the bony growth of the orbit and surrounding ocular structures are also influenced by the changes within the eye.
The ophthalmologist caring for the pediatric patient must understand the normal developmental changes so as not to confuse them with pathologic states. In addition, disease states may interfere with normal development. Intervention must occur during critical periods to allow for normal growth of the eye and orbit. Premature infants can exhibit findings normal in the embryonic development consistent with the infant’s gestational age and not the chronologic age. This chapter will address the normal changes in the globe, anterior segment, pupil, retina, neurologic development, orbit, and refractive status of the eye from findings in premature infants through to complete maturation of these structures.
GLOBE DIMENSIONS
The weight of the term infant eye varies between 2.3 and 3.4 g (1). The average adult eye weighs 7.5 g. The volume of the infant globe varies between 2.20 and 3.25 cm3. The average axial length of the eye in term infants is between 16.8 and 17.5 mm when measured ultrasonographically (2). The value is slightly less when measurements are obtained through histopathologic studies. Normative data reveal a triphasic pattern of increase in the axial length (2). During the first year of life, the rate of growth is the greatest. There is on average a 2.5 to 3.8 mm increase in the axial length in the first year of life, making the mean axial length 20.7 mm. The rate of increase then decreases in subsequent years, such that the mean axial length is 21.5 mm in the second year and 21.9 mm in the third year. Thereafter, the rate of growth slows to approximately 0.4 mm/year. The axial length generally reaches adult dimensions by approximately 5 years of age. Between 5 and 15 years of age, there may be small increases in the axial length of usually <1.0 mm without the presence of myopic refractive error. The mean axial length in girls is shorter than boys through adolescence, 23.92 and 24.36 mm, respectively (1).
In pathologic conditions such as congenital glaucoma, cataracts, and retinopathy of prematurity, the axial length measurements vary from these normative values. In a group of 170 children with congenital and infantile cataracts, mean axial length was 17.86 mm between 0 and 3 months of age and 21.96 mm between 30 and 42 months of age (3). The range of axial length in this cohort was 14.22 to 25.98 mm. Another study reported shorter axial lengths in the first year of life for cataractous eyes compared with noncataractous eyes—mean axial lengths of 17.9 and 19.2 mm, respectively (4). The standard deviation in mean axial length in children with cataracts was twice the value in noncataractous eyes. The change in axial length may also vary in children with cataracts. Trivedi reported a 0.19 mm/month change from 6 to 18 months and 0.01 mm/month change from 18 months to 18 years of age (4). In addition, African-American children in the study had statistically significant longer axial lengths than Caucasian children (4). The selection of appropriate intraocular lens correction remains a controversial subject. In children with prematurity, there is greater variability in the axial length compared with age-matched controls (5). Increased axial length in children with retinopathy of prematurity contributed to severe myopia, but changes in refractive parameters in the anterior segment were also significant (5). Finally, measuring axial length changes allows for monitoring of control in congenital glaucoma.
Cornea
The absolute dimensions in the term “newborn eye” are closer to the adult dimensions than nearly any other organ in the body. The cornea undergoes macroscopic and intracellular changes to allow transparency, as well as changes in refractive power postnatally, yet the corneal diameter undergoes only minimal growth. The cornea begins with equal sagittal and transverse diameters. In premature infants, the corneal diameter may be approximated at any gestational age because of its relationship to the child’s weight in grams. The corneal diameter in mm equals 0.0014 (weight in grams) plus 6.3 (6). At term, the average corneal diameter horizontally is 9.0 to 10.5 mm, with a mean of 9.8 mm. The vertical diameter may exceed the horizontal with a range of 9.9 to 10.5 mm. In general, a macrocornea is defined as having a horizontal diameter >2 SD from the mean or 11.0 mm in term infants. A microcornea has a diameter < 9.0 mm (Fig. 2.1). This standard range of measurements may help identify children with corneal enlargement secondary to diseases such as infantile glaucoma, once racial variation has been taken into account. The range in corneal diameter was slightly greater in a cohort of African babies, 9.0 to 12.5 mm at 1 week of life, showcasing the importance of evaluating normative data in regard to ethnicity (7).
The growth in corneal diameter is also accompanied by changes in corneal curvature. Since the cornea is instrumental in refraction, the changes in corneal radii of curvature influence the clarity of retinal images. To maintain emmetropia, changes in corneal curvature must be perfectly balanced with changes in the lens and axial length of the eye. The corneal curvature is much steeper in infants than in adults. This observation extends to premature infants as well. In premature infants with a mean gestational age of <32 weeks, the mean keratometry was 63.3 ± 3.3 D in the horizontal meridian and 57.3 ± 2.6 D in the vertical meridian (8). These values decreased rapidly to 54.0 ± 3.0 D and 50.7 ± 2.4 D, respectively, by 36 weeks of gestational age. The corneal curvature continues to decrease rapidly in the first 2 to 4 weeks of life in term infants and then slows after 8 weeks of life. Keratometry values obtained for term infants range from 48.06 to 47.00 D. In one study of 4,881 children aged 6 to 14, keratometry values in the horizontal median remained stable through childhood, but the vertical meridian power decreased slightly with age (9). Girls had statistically significant greater power in the vertical and horizontal meridian at all ages compared to boys: 44.27 D compared to 43.52 D vertically and 43.17 D compared to 42.49 D horizontally, respectively. Asian children had greater corneal power in the vertical meridian compared to other ethnicities, and white children had greater corneal power in the horizontal meridian. Ethnicity must be taken into account in the interpretation of normative data.
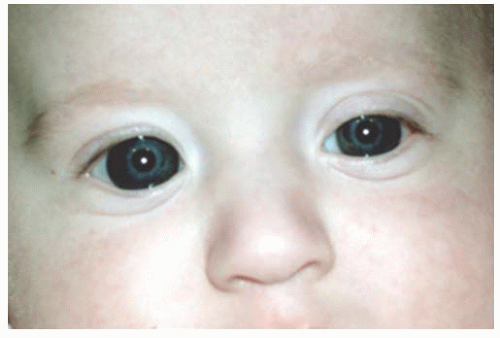 FIGURE 2.1. Megalocornea. The right corneal diameter is 14.0 mm and the left horizontal corneal diameter is 12.0 mm. Each cornea is clear, and there is no evidence of glaucoma. Megalocornea is usually a bilateral condition with corneal horizontal diameter >2 SD from the mean or 11.0 mm in term infants. |
The flattening of the cornea persists into the second and early third decades (10). The average keratometry at age 20 is 42.0 D. The horizontal meridian begins to steepen between the fourth and fifth decades of life and then continues to steepen with age. This results in a gradual change from with-the-rule astigmatism common in youth to againstthe-rule astigmatism in 50- to 60-year-old individuals (10). Advances in corneal tomography have lead to greater precision in measuring the cornea in childhood (11).
Histologically, the layers of the cornea develop to attain both the structure and the function needed in mature corneas. The corneal epithelium thickens with successive cellular layers, and there is an increase in cell size within each layer. At 20 weeks’ gestational age, the corneal epithelium has only two cellular layers, with the basal cells having a thickness of 20 µm (12). By 6 months of age, the basal cells reach their adult thickness of 18 µm. The desmosomes, which are the intracellular junctions between the corneal epithelial cells, are present in a 20-week gestational-age cornea. They are more abundant in the superficial layer, although thinner and less regularly distributed. Corneal abrasions in the early gestational period would, therefore, be expected to occur more easily. In a study of 1- to 12-week-old children, presenting for well visits, 49% had asymptomatic corneal abrasions (13). Most abrasions in this group resolved within 24 hours. The epithelial basement membrane becomes thicker and more homogenous during the early gestational period. The type IV collagen composition of the infant epithelial basement membrane varies significantly from the adult counterpart (14). The hemidesmosomes anchoring the basal cell layer of the epithelium to the basement membrane increase, and the Bowman’s layer becomes compact with more collagen fibrils.
The corneal stroma also becomes thicker during the first several months of life, from 0.229 mm at 20 weeks’ gestation to 0.490 mm at 6 months postnatally, at which time the adult thickness has essentially been reached (12). This change in the thickness results from enlargement of the
collagen fibrils themselves. Once mature, the collagen fibers no longer thicken during subsequent aging. The average diameter of a collagen fiber is between 250 and 300 Å. Most studies report a slow increase in central corneal thickness until approximately 10 years of age (15,16), although some studies have reported no change (17,18). Some of this discrepancy results from the method of measurement used for central corneal thickness, with specular microscopy yielding lower values than ultrasound pachymetry.
collagen fibrils themselves. Once mature, the collagen fibers no longer thicken during subsequent aging. The average diameter of a collagen fiber is between 250 and 300 Å. Most studies report a slow increase in central corneal thickness until approximately 10 years of age (15,16), although some studies have reported no change (17,18). Some of this discrepancy results from the method of measurement used for central corneal thickness, with specular microscopy yielding lower values than ultrasound pachymetry.
There are small structural changes observed between adolescents’ collagen fibers and those from elderly individuals. Adolescent corneal stroma has increased interfibrillary distance compared with samples from elderly individuals. The etiology of the increased cross-sectional area within the corneal collagen fibers has also been investigated. On average, the cross-sectional area increases from approximately 3.04 to 3.46 nm2 over a 90-year timespan. In contrast, the interfibrillary distance decreases with age. It has been postulated that this change may be due in part to an increase in the nonenzymatic cross-linking between collagen molecules. Biochemical studies have revealed an increase in collagen glycation and its end products within elderly corneal stromal samples resulting in decreased interfibrillary spacing. Counterbalancing the decrease in interfibrillary distance, there is an increased occurrence of water accumulation between collagen fibrils. These “lakes” contribute to light scatter, which may diminish visual clarity with age, and largely account for the overall increased cross-sectional area.
The keratocytes in the corneal stroma decrease in thickness and density during development. One study found a keratocyte density of 6.22 × 104 keratocytes/mm3 in the first decade of life, decreasing approximately 0.3% per year thereafter (19). This study also found that interindividual keratocyte density was quite variable, while intraindividual density was not. Keratocytes play an important role in corneal stromal wound healing. It has been postulated that the decrease in keratocyte density is due to a combination of environmental and predetermined genetic factors, although the exact mechanism is unknown. The decrease in keratocytes may be partially or completely responsible for the age-related changes within the cornea, including the decrease in central corneal thickness with age, steepening of the cornea with resulting refractive changes through childhood, and increased light scatter noted in the cornea with age. The decline in keratocyte density and the interindividual variability may be of particular importance to refractive procedures where corneal stromal wound healing may affect the outcome of the procedure. An inverse relationship has been noted between the vigor of wound healing with its effect on refractive regression and increasing age.
Like corneal keratocyte density, endothelial cell density also decreases with age. A cornea from a 12-week fetus has an endothelial cell density of 14,000 cells/mm2 (19). At term, the average endothelial cell density is 6,800 cells/mm2. This large decline during fetal development may be explained by rapid corneal growth. Yet, the decrease from infancy to childhood is also rapid, ranging from 1.4% to 4.0%. As mentioned previously, corneal growth has essentially attained adult parameters by 2 years of age. The reason for the further decrease in endothelial cell density is unknown. The annual rate of loss of endothelial cells slows in adulthood to approximately 0.3% per year. This rate of decrease emphasizes the importance of approximate age matching of donor corneas used in corneal transplants in newborns and infants with conditions of visually limiting corneal opacities.
The endothelial cells contribute to the translucent nature of the cornea as well as to the regular spacing of the collagen fibers. The cornea is an intransparent structure in fetuses up to 26 weeks’ gestational age (20). The intransparency is mild to moderate, symmetric, and uniform with smooth corneal epithelium. The cornea becomes transparent within 4 to 6 weeks of birth (Fig. 2.2). Most infants >32 weeks’ gestational age demonstrate corneal transparency, but any developmental intransparency clears within 1 to 2 days of delivery in term infants.
The etiology for the variability in the structural changes in the cornea has long been sought. Recent studies have identified proteins that may mediate cellular activities in corneal development (21). Tenascin-C (TN-C) mediates several important cellular activities, including cell adhesion, migration, and proliferation and differentiation of stem cells. TN-C is expressed widely in the preterm cornea. TN-C is expressed on both surfaces of Descemet membrane in infant corneas (22). Restriction of expression begins to occur in the neonate, and by adulthood TN-C is expressed only in the limbus. Variants of TN-C caused by alternate splicing of the gene lead to the pleiotropic nature of TN-C. These variants also differ in their expression within the cornea and with age. Rho-mediated signaling in injury can induce expression of TN-C in adult corneas (23). Further studies of proteins that influence growth may lead to development of strategies to effect disease states.
In summary, the corneal diameter increases slightly, especially in the vertical diameter, and the corneal curvature flattens in childhood. Early in the second decade of life,
the corneal curvature steepens, especially in the horizontal meridian. The corneal layers also mature in the first decade. There is an increase in the size of the epithelial and stromal layers in the early postnatal period, but the endothelial and stromal keratocyte density decreases in the same time period. Nevertheless, there is an increase in the overall corneal thickness.
the corneal curvature steepens, especially in the horizontal meridian. The corneal layers also mature in the first decade. There is an increase in the size of the epithelial and stromal layers in the early postnatal period, but the endothelial and stromal keratocyte density decreases in the same time period. Nevertheless, there is an increase in the overall corneal thickness.
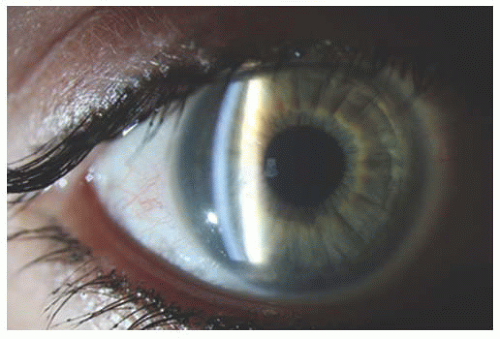 FIGURE 2.2. Slit lamp exam of cornea revealing slight opacity in cornea consistent with very mild congenital hereditary endothelial dystrophy. |
Corneal thickness influences the measurement of intraocular pressure (24,25). Intraocular pressure measurement may be overestimated as corneal thickness increases. The positive correlation in central corneal thickness and intraocular pressure is generally established in adult populations (26). By extrapolation, thinner corneas in infants <6 months of age would be expected to produce lower intraocular pressures with Goldmann or Schiotz tonometry. Furthermore, the correlation of central corneal thickness and intraocular pressure in children is variable due to variation in central corneal thickness with age and race (18). One study found intraocular pressure varies with age according to the relationship, Ta equals 0.71(age in years) plus 10, until the age of 10 (27). The method of intraocular pressure determination also significantly impacts the value of the intraocular pressure (28). Applanation tonometry underestimates intraocular pressure under general anesthesia and generally underestimates intraocular pressure in childhood according to some studies (27,29). General anesthetics tend to lower intraocular pressure, while infant distress with crying and squeezing of the lids elevates intraocular pressure. Since the Tono-Pen compresses a much smaller corneal area, it is slightly less affected by corneal thickness (30). Some studies have found that intraocular pressure increases 2.1 to 3.5 mmHg with every 100 µm increase in central corneal thickness in children when measured with the Tono-Pen (18,31). These variations in intraocular pressure with corneal characteristics must be incorporated into the management of infantile glaucoma.
Anterior Chamber
The anterior chamber depth is influenced by the growth of the sclera, as well as by factors related to lens movement and thickness. There are many methods to evaluate the anterior chamber including slit-lamp photography, ultrasound biomicroscopy, and anterior segment optical coherence tomography (32). Using some of these methods, estimates of the anterior chamber depth average 2.05 mm with a range of 1.8 to 2.4 mm in depth (33). The depth continues to increase until the end of adolescence, and then it progressively diminishes. In emmetropic patients, the increase in anterior chamber depth appears to stop at an earlier age, compared to patients with myopia (34). This apparent difference is related to the continued changes in the lens and axial length in patients with myopia. The anterior chamber depth appears to vary with ethnicity as well (9). Native American children had the least change in anterior chamber depth from ages 6 to 14 with an average depth of 3.50 mm. African-American children had the greatest change during this same period, from 3.41 to 3.62 mm. After 12 years of age, the anterior chamber depth appears to be constant regardless of ethnicity.
Care must be taken with the measurement technique as with physiologic accommodation, the anterior chamber depth diminishes approximately 24 µm/D using slit-lamp adapted optical coherence tomography (35). After instillation of mydriatics, the anterior chamber depth may increase (36,37). The difference in the anterior chamber depth between the two eyes does not exceed 0.15 mm in normal individuals (34). The anterior chamber depth is slightly deeper in boys than girls, 3.64 mm compared to 3.56 mm, respectively (9). The volume of the anterior chamber is approximately 64 mm3 in term infants and 116 mm3 in adults (34). Adjustments during intraocular surgery in childhood may be necessary based upon these differences in anterior chamber depth.
Iris
The architectural crypts of the iris develop from gestation through the early postnatal period. The primary papillary membrane forms early in gestation and atrophies near term. The color of the iris results from pigmentation of the stromal mesodermal cells and iris blood vessels. At term, the mesodermal stromal cells of the iris continue to develop pigment, which accounts for the darkening of iris color observed in the first few months of life. Researchers have identified many pigment-associated genes that reside on chromosome 15, which contribute to iris color (38). One particular sequence within the regulating element of the HERC2 segment on chromosome 15 accounts for 74% of the variance in human eye color (39). The contour of the iris including Fuchs’ crypts, contraction furrows, Wolfflin nodules, and nevi influences the perceived color of the human iris. Separate genes regulate these iris patterns, including SEMA3A and TRAF3IP1, which are also associated with pathways that control neuronal pattern development (40).
Lens
The tunica vasculosa lentis is a plexus of blood vessels that is instrumental in the development and nourishment of the lens in embryonic life. The tunica vasculosa lentis completely regresses after 35 weeks of gestation. Although the exact cellular processes involved in the involution of hyaloid vessels remain unclear, autophagy is thought to play a role (41). The extent of regression can be used to estimate gestation age postnatally as well (42). At 27 to 28 weeks’ gestational age, the entire lens surface is covered with vessels. Between 29 and 30 weeks, the central vessels of the tunica begin to atrophy. At 31 to 32 weeks, the central lens is visible, with thinning of the peripheral vessels. Between 33 and 34 weeks of gestation, only thin peripheral vessels remain of the tunica vasculosa lentis.
The crystalline lens is the structure most responsible for adapting to the changing axial length of the eye and its subsequent influence on the refractive needs of the eye. The length of the eye increases rapidly until approximately 3 years of age, followed by slow growth of approximately 1 mm in the next 10 years. The cornea loses approximately 3 to 5 D of power by flattening in the first year of life, leaving the majority of the dioptric change necessary to maintain emmetropia to the lens. A tremendous decrease in dioptric power occurs in the first year of life. The eye’s power changes from approximately 90 D at birth to 75 D at 1 year. Despite the axial growth that necessitates this change, the majority of infants maintain emmetropia.
Several authors have attempted to study the structural, molecular, and geometric changes in the lens that allow it to change in power (43,44,45). New epithelial cells located adjacent to the lens capsule elongate and differentiate into fiber cells throughout life. These cells produce the beta- and gamma-crystallins that make up the body of the lens. These fibers congregate in the nuclear region of the lens with increasing concentration of protein, which increases the refractive index of the lens. Lens growth occurs in two phases. Prenatally, there is a sigmoidal rapid growth rate, which generates approximately 149 mg of the lens tissue (45). This prenatal growth generates the adult nuclear core of the lens (46). Growth continues throughout life, but the growth rate becomes linear after 6 to 9 months of age, approximately 1.38 mg/year (45). There are no gender differences in the rate of growth of the lens, and the lens thickness does not differ between boys and girls regardless of age (9). There is thinning of the lens during the first three years of life due to the equatorial growth of the eye, which essentially stretches the lens (47). Consistent with this change, the anterior and posterior lens radii increase in childhood by 1.0 and 0.2 mm, respectively (44). Equatorial growth would cause passive stretching of the crystalline lens with flattening of the lens surface curvature and reduction of the lens power. Interestingly, the rates of increase of the anterior and posterior lens radii differ. The rate of increase of the anterior lens curvature slows after age 3, whereas the posterior lens curvature rate remains constant throughout childhood. These both contribute to the overall flattening of the lens. At age 6, the lens thickness varies between 3.50 and 3.60 mm depending on ethnicity (9). Hispanic children had the thinnest lenses and African-American children had the greatest thickness. In all ethnicities, the lens decreased in thickness until age 11 to 12 and then began to gradually increase in thickness.
The composition of the lens undergoes molecular changes as well. Fetal lenses have a higher percentage of gamma crystalline protein (21%) compared to adolescents (13%). Gamma crystalline proteins have excellent solubility and stability that prevent the scatter of light (48). The beta and alpha crystalline percentages are similar in childhood. In the elderly on the other hand, the alpha crystalline proportion is greater. The optical density of the lens also increases throughout life, increasing the absorption of light (49). Many proteins play a role in lens formation and in differentiation of lens cells. One of the factors found to influence lens morphology is Epha2. Epha2 is expressed by the epithelial cells of the lens and plays a role in directing lens fibers to the correct location. In the absence of Epha2, fiber cells migrated off the optical axis into new suture lines (50). Further studies will continue to elucidate the proteins involved in lens maturation leading to the possibility of influencing those pathways in the setting of pediatric cataracts.
Sclera
The sclera is predominantly an extracellular collagenous matrix. In adult eyes, the thickness varies from 0.53 ± 0.14 mm at the limbus, 0.39 ± 0.17 mm at the equator, and finally approximately 1.0 mm near the optic nerve (51). The total surface area is 16.3 ± 1.8 cm2. The collagen in the sclera undergoes developmental changes in the early postnatal period. The sclera is four times as pliable in infants as in adults and has approximately one-half the tensile strength. This pliability explains the buphthalmos seen in infantile glaucoma with elevated intraocular pressure.
The structural changes in the sclera are due to the changing proteoglycan composition of the sclera. There are three major proteoglycans in the sclera: aggrecan, biglycan, and decorin (52). All three are increasingly expressed until the fourth decade. There is an increase in sclera thickness from 0.45 mm in neonates to 1.09 mm in adults. After the fourth decade, decorin and biglycan decrease in expression. Aggrecan has the highest concentration in the posterior sclera and continues to show high expression throughout life. The different expression rates of these proteoglycans may result in the differential growth seen in various portions of the sclera (52). For example, growth of the posterior sclera may result in increases in axial length, which will be discussed in more detail in the refractive section. Certain anatomic relationships confirm the differing growth rates. The posterior portion of the sclera shows greater growth than the equatorial portion in the early postnatal period, resulting in the apparent forward migration of the extraocular muscle insertion sites relative to the equator of the eye (53).
In a primate model, the sclera thinned in older monkeys and became more structurally stiff (54). Collagen fibers with increased stiffness oriented circumferentially around the optic nerve may contribute to the greater sensitivity of nerve fibers to intraocular pressure changes. The sclera also becomes less permeable with age (55). These structural changes arise from changes in the collagen and proteins that compose the sclera. Lumican is a keratin sulfate proteoglycan in the sclera that appears to regulate collagen fiber formation and organization. As the sclera ages, lumican interacts with aggrecan, forming a complex that becomes more abundant with age (56). In addition, glycation end products and advanced lipoxidation end products increase in the sclera with age (57).



