(1)
University of Sydney, Sydney, Australia
Overview
1.
Mechanisms that broaden our luminance range for vision
Our visual system can function over a wide range of light intensities, from starlight to a bright sunny day – a luminance range of 10 10 (10 log units) [1].
Several dynamic mechanisms exist that broaden our luminance range for vision in response to a change in ambient level of illumination.
They allow the visual system to obtain maximal visual information at each luminance level:
(a)
Light-induced changes in pupil size
(b)
The switch between the scotopic and photopic pathways in our duplex visual system
(c)
Visual adaptation
2.
Visual adaptation: change in gain of the visual system
Adaptation is an alteration in gain of the visual system.
Gain is the ratio of the output signal (neural responses) to input signal (light).
Gain increases under dim conditions and decreases under bright conditions.
Visual adaptation is largely mediated by:
(a)
Retinal photoreceptor mechanisms
(b)
Retinal neural channel mechanisms
(c)
Higher center mechanisms
Each mechanism is involved in light adapation and dark adaptation.
3.
Light adaptation
As background luminance increases, light adaptation processes maximize spatial, temporal, and chromatic contrast resolution.
This allows the visual system to make complex discrimintions such as contour detection, fine spatial resolution, movement, and color perception.
However, there is a corresponding decrease in sensitivity. For example, the dark-adapted eye can see stars at night; during daylight (photopic conditions), the stars are equally bright but not seen.
4.
Dark adaptation
Dark adaptation is the ability of the visual system to recover sensitivity following light exposure.
Compared to light adaptation, dark adaptation is a slower process.
Most dark adaptation occurs within the first few minutes however takes more than 30 min to complete (see Fig. 21.1a) [5].
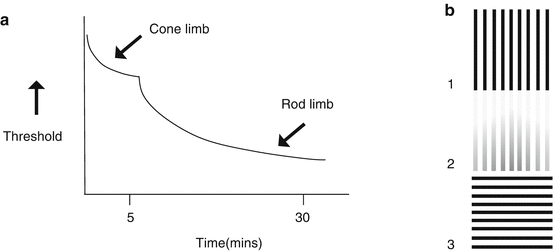
Fig. 21.1
(a) The dark adaption curve (based on Hecht et al. [5]). (b) Perceptual reduction in apparent contrast (Adapted from Kohn [6]). Staring at the vertical bars in 1 for 30 s reduces the ability to detect a low-contrast portion (the top) of image 2. The horizontal bars in 3 produce a less strong adaptative effect
5.
Contrast adaptation
Contrast adaptation affects our visual ability to discern spatial and temporal contrast of stimuli.
Unlike light and dark adaptation, it is not influenced by changes in ambient light levels, unless a shift from photopic to scotopic range occurs.
The strength of adaptation is related to the similarity between the adapting and test stimuli (see Fig. 21.1b) [10, 11].
It occurs in the visual cortex, lateral geniculate nucleus (LGN), and inner retina [6].
Mechanisms for Broadening the Dynamic Luminance Range of Vision (Table 21.1)
Mechanism | Overview | Sensitivity range (log units) | Time from stimulus to adaptation |
|---|---|---|---|
Pupil size | Pupil size reduces in bright light and increases in dark, modulating light entering the eye It is mediated by the pupillary light reflex | 1.2 | 1 s |
Switch from scotopic to photopic systems | Scotopic vision facilitates light detection in dim light Scotopic vision is mediated by rods and their retinal neural channels Photopic vision facilitates contrast, color, and motion descrimination in medium to bright light Photopic vision is mediated by cones and their retinal neural channels | Each system can operate over 4–5 log units with some overlap (1–2 log units) | Milliseconds |
Visual adaptation | |||
A. Photoreceptor mechanisms | Rod responses are easily saturated by increased ambient light intensity. The range of scotopic sensitivity is greatly enhanced by post-receptoral neural channels Cones escape saturation no matter how intense the steady light Light-induced changes responsible for adaptation include: 1. Pigment bleaching and regeneration 2. Alterations in intracellular Ca2+ levels 3. Alterations in phosphodiesterase activity | Rods: 1–2 Cones: 5+ | Light adaptation: 1. Cones: milliseconds 2. Rods: slower than cones, <1 s Dark adaptation: Rate limited by pigment regeneration 1. Cones: 3–5 min 2. Rods: 10–30+ min |
B. Retinal neural mechanisms | Light and contrast adaptation processes occur in retinal neural channels Mechanisms include: 1. Electrical coupling 2. Lateral inhibition 3. Ganglion cell adaptation to signal | 3 | Milliseconds – minutes |
C. Higher visual center mechanisms | Higher visual center neurons are capable of contrast but not light adaptation Contrast adaptation has been demonstrated in the lateral geniculate nucleus magnocellular layers and cortical areas V1, V2, and MT/V5 The strength of adaptation is related to the similarity between the adapting stimulus and test stimulus Mechanisms include: 1. Hyperpolarization of the cell-soma membrane 2. Presynaptic depletion of glutamate 3. Modulation of neural responses by activity of neighboring neurons | N/A | Milliseconds – minutes |
1.
Change in pupil size
The pupil size enlarges in the dark to 8 mm and constricts in light conditions to 2.5 mm.
This corresponds to 1.2 log units of luminance range.
Although a relatively small contribution to the dynamic luminance range, changes in pupil size provide rapid dynamic shift in light or dark while other adaptive processes are taking place.
2.
The duplex system: switching from scotopic to photopic states
Each system includes photoreceptors and their retinal neural processing channels.
Scotopic and photopic vision vary in fundamental ways (Table 21.2) [48–52, 57–64].
Scotopic
Photopic
Photoreceptor type
Rod
Cone
Background luminance
Low
Medium – high
Luminance range (log units)
−4 to −1
1–4
Maximum spectral sensitivitya
507 nm
555 nm
Color vision
Absent
Present
Spatial resolution
Poor
Good
Spatial summation
Increased
Decreased
Increment luminance sensitivity
High
Low
Contrast sensitivity
Low
High
Site of maximal acuity
7° from fovea
Fovea
Foveal scotoma
Present
Absent
Temporal resolution
Poor
Good
Critical duration (Tc)b
Long
Short
The rod system allows maximal light detection sensitivity in scotopic conditions, with high gain at the expense of temporal and spatial acuity.
The cone system provides maximal temporal and spatial acuity in photopic conditions with low gain at the expense of sensitivity.
In modern urban life, the majority of our vision uses the photopic system; only in exceptionally dark conditions (e.g., starlight, dark rooms) do we rely on the scotopic system [3].
As background light intensity shifts from low to high luminance levels, so does our reliance from the rod to cone systems.
Mesopic conditions are intermediate between scotopic and photopic, such as a moonlight night; vision in these conditions is mediated by interaction between the rod and cone systems [15].
3.
Photoreceptor mechanisms of visual adaptation
(See “Photoadaptation in rods and cones” in Chap. 8, The Retina)
The magnitude and speed of photoreceptor membrane potential responses to light stimuli are influenced by background luminance levels [21].
Significant post-receptoral changes extend the scotopic system’s range beyond that of rods [18].
Mechanisms include:
(i)
Visual pigment bleaching and regeneration
Photoreceptor pigment is rapidly bleached on bright light exposure, resulting in separation of the chromophore from opsin (see “The phototransduction cascade” in Chaps. 8, The Retina, and 9, The Retinal Pigment Epithelium).
Free opsin activates transducin directly, although less powerfully than metarhodopsin II.
The decay of photopigment in bright light reduces the magnitude of the photoreceptor response to light, resulting in light adaptation.
This results in less free opsin, increased photoreceptor pigment available for light detection and increased light sensitivity.
(ii)
Light–induced reductions in cytoplasmic Ca 2+ levels
It causes modulation of the cationic nucelotide-gated (CNG) channels reducing light sensitivity.
(iii)
Increased photodiesterase activity in steady light
This results in more rapid turnover of cGMP, reducing light sensitivity [22].
4.
Neural adaptation
Neural adaptation mechanisms include retinal and higher visual pathway processes.
They provide 1000× (3 log units) of adaptative range.
These processes are very rapid (occuring in milliseconds).
In the light, they decrease spatial and temporal summation (causing less efficient light detection) and increase surround inhibitory effects (providing more efficient contrast discrimination).
5.
Retinal neural adaptation mechanisms
(i)
Electrical coupling of photoreceptor, horizontal, bipolar, and amacrine cells [12].
Electrical rod–rod coupling is important in dark adaptation.
It spatially averages rod signals over large distances, which
(a)
Decreases noise filtering at the rod-bipolar junction
(b)
Increases rod synaptic saturation
This improves light detection at the expense of image resolution [12].
Rod–cone coupling encourages shift to the photopic range enhancing light adaptation [17]. Additionally, it allows maximal rod responses to light to reduce cone sensitivity [16].
(ii)
6.
Adaptation in higher visual areas
Get Clinical Tree app for offline access

Spatial and temporal contrast adaptation has been demonstrated in LGN magnocellular neurons, primary visual cortex (V1), and extrastriate cortical areas V2 and MT/V5 [6, 10, 11, 36, 38].
Adaptation is strongest when the adapting stimulus closely resembles the test stimulus.
This is because adaptation alters the sensitivity of individual cortical neurons that are tuned to specific spatial frequencies and orientations.
Mechanisms include:
(i)
Hyperpolarization of the cell-soma membrane
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree

