, Caroline Huart1, 2, Basile Landis4, 5 and Thomas Hummel4
(1)
Department of Otorhinolaryngology, Clinique Universitaires Saint-Luc, Brussels, Belgium
(2)
Institute of Neuroscience, Université catholique de Louvain, Brussels, Belgium
(3)
HNS & ENT Department, Cliniques Universitaires Saint Luc, Brussels, Belgium
(4)
Department of Otorhinolaryngology, Smell and Taste Clinic, Clinic of the Technical University of Dresden Medical School, Fetscherstrasse 74, Dresden, 01307, Germany
(5)
University of Bern Swiss, Berne, Switzerland
Abstract
Intranasal trigeminal system interacts with the olfactory system to provide a complete chemosensory perception of the odorants stimuli. The perception of nasal patency and nasal airflow is also mediated by trigeminal afferents. The trigeminal system is therefore mandatory for both the chemosensory and the somatosensory perception and nasal mucosa with trigeminal nerve endings needs to be preserved in order to assume these two functions.
Psychophysical testing of intranasal trigeminal function has not yet been established in clinical routine. However, in some specialized centers of for research purpose, electrophysiological recordings (negative mucosal potential and trigeminal event-related potentials) and functional imaging may be obtained.
Finally, the intranasal trigeminal system is also capable of inducing a neurogenic inflammation mainly through an axon reflex located in the subepithelial level of the nasal mucosa.
Keywords
Chemosensory functionTrigeminalOlfactionReflexesSomatosensory functionCore Messages
Intranasal trigeminal system mediates the sensation of temperature, pressure, perception of nasal airflow during breathing, nociception and participates to the chemosensory perception of odorant stimuli.
Chemosensory perception is not only mediated by free nerve endings in the nasal mucosa but also by some trigeminal fibers in close contact with solitary chemosensory cells.
Besides the sensory nerves, the parasympathetic and the orthosympathetic systems play an important role in the normal physiology of the nose
Testing the intranasal trigeminal function, both psychophysically and electrophysiologically, is possible and may be used in the assessment of a patient with a chemosensory dysfunction.
Healthy subjects need to have intact trigeminal and olfactory systems to have a full complete picture of the chemosensory stimulus.
Olfactory and trigeminal systems interact both at a central and peripheral level.
In patients with olfactory loss, a compensatory mechanism probably exists between the olfactory and the trigeminal systems.
17.1 Introduction
The nasal mucosa through the intranasal trigeminal nerve is a full sensory organ, functionally organized and responsible for both the nasal patency perception and the chemosensory perception and also responsible to a certain degree for nasal inflammation. The primary function of the intranasal trigeminal system is to protect the upper and lower airways for potential life-threatening substances acting as a sentinel to shorten or stop inspiration reflexively.
Besides this protective somatosensory function, the intranasal trigeminal system also helps to the global chemosensory perception with the olfactory system. Indeed, most of the odorants stimulate the neural olfactory and intranasal trigeminal systems (Doty 1995).
Finally, the intranasal trigeminal system is also capable of inducing a neurogenic inflammation mainly through an axon reflex located in the subepithelial level of the nasal mucosa.
The olfactory (cranial nerve I) and the trigeminal (cranial nerve V) systems interact at different levels and this interaction is essential for the odor sensation (Cain and Murphy 1980). The olfactory system is more dedicated in identification task for hedonicity and alimentary behavioral, recognition and memory, behavioral and social comportments than the trigeminal system probably more oriented to protective function and reflexes.
17.2 The Nerves of the Nose
Sensory nerve endings from branches of the trigeminal nerve are located in the epithelia of the nose and sinuses, the eyelids and the cornea, the oral cavity and the skin. Fibers from the intranasal trigeminal nerve mediate the tactile sensation of temperature, pressure and perception of nasal airflow during breathing and participate to the chemosensory perception of odorant stimuli. Trigeminal receptors are located throughout the epithelia of the nasal mucosa and contribute to global perception of odorous stimuli reaching the nasal fossa and the upper airway.
The nasal cavity is innervated by two branches of the trigeminal nerves, i.e., the ophthalmic and the maxillary branches. The ethmoid nerve innervating the anterior nasal mucosa and the external surface of the nasal fossa is part of the ophthalmic division while the nasopalatine nerve which innervates the posterior part of the nasal cavity is part of the maxillary division. The trigeminal nerve has chemosensory and mechanosensory fibers. Mechanosensory fibers are large fast-conducting Aβ-fibers. Thin and fast-conducting myelinated Aδ-fibers and thin and slow-conducting unmyelinated C-fibers are responsible for thermoreception (cold and warm stimuli) and for nociceptive perception (pain, painful mechanical, noxious chemical stimuli). The sensations mediated by the trigeminal nerve are usually described as burning, stinging, itching, tickling, cooling and warming feeling. Trigeminal free nerve endings have receptors which may be activated through several factors such as changes in pressure, temperature, irritants, and humidity. Substance P, calcitonin gene-related peptide (CGRP), and other neuropeptides are found in the trigeminal nerve fibers (Finger et al. 1990). Some trigeminal fibers are in close contact with solitary chemosensory cells located in the nasal epithelium and more responsible for chemosensory perception because they are responsive to both bitter tastants and chemical irritants (Fig. 17.1).
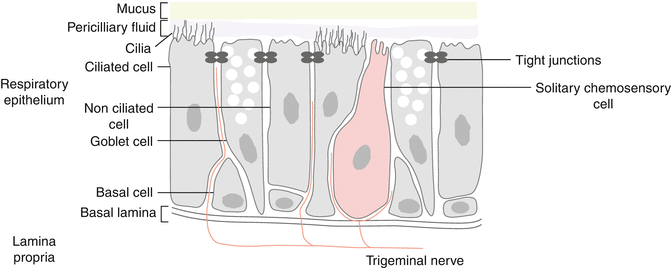
Fig. 17.1
Trigeminal fibers in close contact with solitary chemosensory cells and trigeminal nerve free endings located in the nasal epithelium and responsible for chemosensory and somatosensory perceptions
At the receptor level, one of the first described nociceptors was the ion channel receptor family and characterization of one of these receptors was obtained with nicotinic acetylcholine receptor. Transient receptor potentials (TRP) channels are well expressed on sensory nerves and may influence cell function by mediating the flux of cations across the plasma membrane into the cytoplasm generating action potentials. Ion channels in the TRP family can be opened by many kind of stimuli, i.e., chemical or physical. The TRP family can be subdivided into six subfamilies and many of them are found at the free nerve ending of the trigeminal nerve such as the vanilloid receptor (TRPV1), the purinergic receptor (P2X), the acid sensitive ion channels (ASIC/DRASIC), the channel responsive to menthol (TRPM8) (cooling), the channel responsive to changes in heat and eugenol (TRPV3) (warming), and the channel responsive to isothiocyanate (TRPA1), the major compound of mustard oil (Bessac and Jordt 2008).
Like for the skin sensory perception, the unmyelinated C-fibers (slow conduction) are responsible for burning sensations and the myelinated Aδ-fibers (fast conduction) are responsible for stinging sensations.
The cell bodies of the trigeminal fibers are located in the Gasserian ganglion. Nerve fibers from the cell bodies thereafter participate to the sensory afferent system and project to the trigeminal sensory nucleus that extends from the rostral spinal cord to the midbrain. Interestingly, some individual cells in the ganglion send axons to the olfactory bulb indicating that some interaction exists at this level. Neurons then project to the amygdala and to the ventral posterior medial nuclei of the thalamus. Most of the ascending fibers cross towards the contralateral side with some fibers ascending ipsilaterally (different for the olfactory pathways (Brand 1999)). The nerve projections terminate in the primary somatosensory cortex (SI) and also in the secondary somatosensory cortex (SII) with a right hemispheric predominance (Hari et al. 1997; Rombaux et al. 2008a, b). Trigeminal activation also leads to insular cortex activation and to ventral orbitofrontal cortex mainly to the right side explaining at the central level the interactions with others chemosensory systems like taste and olfaction (Anton and Peppel 1991).
Besides the sensory nerves, the parasympathetic and the orthosympathetic systems play an important role in the normal physiology of the nose (Kaliner 1992). Parasympathetic nerves have acetylcholine as major neurotransmitter and acts on muscarinic receptors to induce increased glandular secretions and vasodilatation. Vasointestinal peptide (VIP) is another neurotransmitter of the parasympathetic system. The sympathetic system with noradrenaline and neuropeptide Y (NPY) as neurotransmitters acts on adrenergic receptors and induces vasoconstriction and increases nasal airway patency (Baraniuk et al. 1991; Baraniuk 1992).
Pathophysiological mechanisms and nasal symptoms are explained by the interdigitation of these neurologic systems, i.e., the trigeminal sensitive afferent (+ efferent axon reflex), the efferent parasympathetic, and the efferent orthosympathetic systems (autonomic systems) (Fig. 17.2).
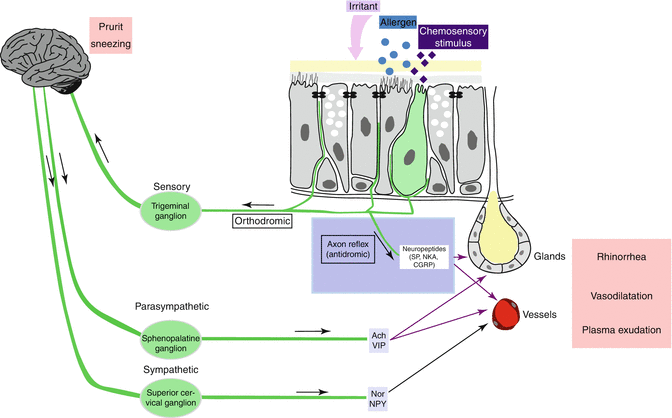
Fig. 17.2
Interdigitation of the neurologic systems found in the nasal mucosa, i.e., the trigeminal sensitive afferent (+ efferent axon reflex), the efferent parasympathetic, and the efferent orthosympathetic systems (autonomic systems)
17.3 Consequences of Activation of Trigeminal Receptor and Nasal Reflexes
The activation of the trigeminal system leads to the perception of potentially noxious stimuli, to a global chemosensory perception of odorant stimuli and to some nasal reflexes.The nasal fossa may be divided into two parts, the anterior one most dedicated to the chemosensory perception and the posterior one most devoted to mechanosensory functions. This has been demonstrated by Frasnelli et al. where it was clearly stated that anterior nasal mucosa is more sensible to chemosensory stimuli than mechanical stimuli, while the posterior nasal mucosa is equally sensible to both chemosensory and mechanical stimuli (Frasnelli et al. 2004). However, thresholds to detect chemosensory stimuli such as CO2 is lower when the stimulus is given in retronasally compared to orthonasally (Melzner et al. 2011).
Therefore, nasal mucosa should not be seen as a homogenous tissue as it exhibits a varying degree of sensitivity to trigeminal stimuli depending on the stimulus quality and location in the nasal fossa (Scheibe et al. 2006).
Activation of trigeminal fibers leads to protective reflexes such as increasing secretions (saliva, tears, nasal mucus), decreasing breathing, sweating initiation, and closure of the nasal passage by augmentation of the turbinate volume.
Trigeminal nerve stimulation also induces many reflexes inducing different responses.The nasal cycle is probably the best known neurologic mechanism leading to a fluctuating congestion-decongestion of the nasal fossa secondary to a changing tone in the vasculature controlled by the autonomic system.
The naso-nasal reflex is supposed to be mediated by the parasympathetic system and explains many exacerbations of rhinorrhea and watery discharge (Baraniuki and Kim 2007).
The naso-ocular reflex is bilateral and mostly contralateral, secondary to chemosensory or tactile or physical stimuli. It induces watery eyes, lacrimation, and redness of the conjunctiva.
The “foot-cooling” reflex is secondary to a cold stimulation at the extremities of the inferior limb inducing in the nose a reduced blood flow and subsequently a nasal decongestion. This is also very similar to the reflexes observed in the nose when cooling of the face induces the same effect. Facial cooling through trigeminal receptors may even induce lower airway symptoms (Koskela and Tukiainen 1995).
The naso-cardiovascular reflex is secondary to trigeminal activation in the nose and is responsible for bradycardia and hypotension, may be present during nose surgery, and is of primary importance for the anesthesiologist.
The naso-respiratory reflex or naso-bronchial reflex is present when cold dry air is presented to the subject’s nose inducing increased lower airway resistance.
Cold dry air stimulus may also be used to induce both long-lasting painful sensations (Lötsch et al. 1998) and secretory response in the nose (Fontanari et al. 1996). This mechanism is thought to be secondary to activation of capsaicin-sensitive fibers; alternatively, the change in the osmotic milieu of the respiratory epithelium may trigger activation of the nociceptive system. This may play a role in the pathophysiology of nasal hypereactivity and in the non-allergic noninfectious group of rhinitis (Bernstein 1991) and would lead to the development of capsaicin-based treatment for the patients suffering from these diseases (Lacroix et al. 1991; Marabini et al. 1991; Stjarne et al. 1991; Blom et al. 1998; Taylor-Clark et al. 2005a, b). Capsaicin delivered intranasally has proven its effect in the treatment of the nasal hyperreactivity found in idiopathic rhinitis patients (Van Rijswijk et al. 2003).
These responses may be present after single presentation of the stimulus or when repeated application of the stimuli is delivered. C-fibers and Aδ-fibers respond differently to repeated chemical stimulus. If stimuli are repeated, the burning painful sensation driven by C-fibers is increased, and this is the contrary for Aδ-fibers giving the stinging sensation. This is secondary to central nervous summation more than increase in the firing of the nerve fibers at the periphery.
17.4 Neurogenic Inflammation
The activation of sensory nerves and the release of neuropeptides from neuroendocrine cells found in the respiratory mucosa with a subsequent neurogenic inflammation may explain at least partially some diseases of the upper and lower airways (Lacroix and Landis 2008).
Stimulation of sensory trigeminal fibers may lead to the release of different neuropeptides such as substance P, neurokinin A (NKA), neuropeptide K (NPK), and calcitonin gene-related peptide (CGRP). These neuropeptides are increased in the upper and lower airways of these patients with airway inflammation in a similar way than the inflammatory components usually described as eosinophils or some proinflammatory cytokines (Shusterman et al. 2003).
There is a strong evidence that neuroendocrine cells, sensory neurons, and proinflammatory immune cells interact and promote inflammation and airway hyperreactivity. Neurotrophins such as nerve growth factor (NGF) or neurotrophins-3–4 are also linked to the development of a neurogenic inflammation.
In animals, dendrites of intranasal trigeminal nerve endings can be stimulated in an antidromic way. This antidromic stimulation is called the “axon reflex” and leads to the release of inflammatory neuropeptides form the varicosities of the nerve, producing vasodilation, increased vascular permeability and glandular activation. This phenomenon has been clinically proven in humans where specific activation of the intranasal trigeminal nerve ending produces nasal obstruction, congestion, watery discharge, and sneezing. This axon reflex probably plays a major role in the development of nasal hyperreactivity, non-allergic noninfectious rhinitis known as idiopathic rhinitis, and even allergic rhinitis via the substance P which exacerbates the eosinophilic recruitment after allergen challenge (for review, see Sarin et al. 2006).
17.5 Psychophysical Testing of the Intranasal Trigeminal Function
Testing of trigeminal function with psychophysics is based on threshold measurement, rating of suprathreshold stimuli, discrimination tasks, and lateralization tasks (Hummel 2000; Frasnelli and Hummel 2005).
Trigeminal function assessed with psychophysical testing revealed that sensitivity decreases with age (Wysocki and Cowart 2003).
Psychophysical evidence exists for qualitative specificity of the human intranasal trigeminal system. The nasal trigeminal system is less sensitive than the olfactory system for the majority of odorant stimuli. Recognition threshold of trigeminal stimulus such as CO2 was measured between 32 and 47 % v/v for stimuli of 200 ms duration at an airflow of 8 l/min at body temperature. The threshold for detection can be lowered if stimulation duration is increased (Melzner et al. 2011).
Considering pain ratings, increase in perceived or painful sensitivity occurs more rapidly for trigeminal stimulus than for olfactory stimuli (Cain et al. 1998).
The trigeminal and the olfactory systems also have a different contribution on the presentation of mixed compounds. In normosmic subjects, trigeminal stimuli are perceived as more intense when they are accompanied by an olfactory stimulus while the olfactory stimulus seems to have no effect when a mixed compound is presented. The trigeminal stimulus may induce an additive or even a hyperadditive effect on the perception after a mixed stimulus presentation (Cornetto-Muniz and Hernandez 1990).
Qualitative discrimination task with trigeminal irritants demonstrate that human are capable to discriminate among different trigeminal stimuli even in the absence of any olfactory stimuli given concomitantly (Laska et al. 1997), even if this ability seems to decrease with age (Laska 2001). In contrast to odor stimulation, trigeminal stimuli can produce increase in pain intensity when repeated stimuli are given with a short interval demonstrating a sensitization effect while on the contrary a desensitization effect exists when repeated stimuli are delivered with long interstimulus interval (Hummel et al. 1994; Brand and Jacquot 2002). Temporal integration of trigeminal informations is thus different than olfactory temporal integration. Psychophysical studies with capsaicin have demonstrated a sensitization effect meaning that the subjective pain rating was increased after the second stimulation if the interstimulus interval was less than 1 min. On the contrary with a second stimulation delivered after 4 min, a desensitization effect was observed. This leads to the idea that repetitive delivery to the nasal mucosa was perhaps a treatment for patients with hyperalgia in the nasal fossa or for patients with non-allergic noninfectious rhinitis (Brand and Jacquot 2002). However, this mechanism is linked to the type of the stimulus and sensitization and desensitization in the nasal cavity do not follow the same processes in relation to the molecules studied (Jacquot et al. 2005).
Lateralization task revealed that trigeminal stimuli are perceived without error when the subjects blindfolded is asked to determine the side of stimulation and that this ability is lost for olfactory stimuli or when the odor has a mixed property between trigeminal and pure olfactory valence (Kobal and Hummel 1990). In others words, pure olfactory stimuli cannot be localized to the nasal cavity while on the contrary pure trigeminal stimuli can be localized. The results are lower in patients with an olfactory dysfunction independent of the cause of the olfactory problem (Hummel et al. 2003).
Subjective ratings of nasal patency are also influenced by the trigeminal system. For example, stimulation of the nasal fossa with menthol is accompanied by an increase of perceived nasal patency (Eccles et al. 1989) while on the contrary anesthesia of the nasal mucosa leads to a perception of decreased nasal patency even in both cases objective nasal patency did not change.
Many studies have been conducted on anosmic subject and trigeminal thresholds were found to be higher in anosmic subjects than in control (Cornetto-Muniz and Cain 1998). Age-related decline of intranasal sensitivity was also reported with psychophysical but also electrophysiological evidence (Frasnelli and Hummel 2003).
17.6 Electrophysiology and Functional Imaging
Electrophysiological recordings from the intranasal trigeminal system may be obtained at the peripheral level, i.e., the negative mucosal potential (NMP) and at the central level by recording cortical responses after delivery of an intranasal trigeminal stimulus, i.e., the trigeminal event-related potential (Trigeminal ERP).
The NMP is recorded from the nasal mucosa and is thought to represent the summated receptor potentials of chemical nociceptors in a very similar way to the electro-olfactogram which represents the global activity of olfactory receptor neurons located in the olfactory neuroepithelium (Thürauf et al. 1991).
Human NMP may be obtained after CO2 intranasal stimulation and the amplitude of the NMP is well correlated with the subjective pain rating (Kobal 1985; Hummel et al. 1996a, b). NMP may also be recorded by stimulating polymodal nociceptors such as TRPA1, TRPV1 and 2. Responses to the NMP are different according to the stimulus used, i.e., CO2, menthol or ethanol (Meusel et al. 2010) and decrease in response to repetitive stimulation (Hummel et al. 1996a, b).
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


