31
Genetics of Age-Related Macular Degeneration
Mike Grassi  Josephine Hoh
Josephine Hoh
Ophthalmology has a rich tradition of providing major contributions to the field of genetics. For instance, the first inherited disease to be linked to a human autosome was the pulverulent cataract, which was linked to the Duffy blood group antigen by Renwick and Lawler in 1963 (1). The first tumor suppressor gene to be discovered, which was also one of the first positionally cloned genes, was the retinoblastoma gene (2). More recently, the implication of the complement factor H (CFH) gene in age-related macular degeneration (AMD) resulted from the first successful genome-wide association (GWA) study for a complex trait (3, 4, 5).
Support for a genetic component to AMD has grown slowly and was initially based on conjecture and anecdotal observation. AMD was initially referred to as “senile macular degeneration” because of the mistaken belief that all elderly individuals would eventually develop this condition and that there was no underlying genetic component. In 1973, Gass et al. published a retrospective case series reporting a 20% prevalence of central vision loss in family members of patients with AMD (6). A more sophisticated epidemiologic approach, the familial aggregation study, was later employed to compare the frequency of AMD in siblings and children of affected individuals to the frequency of AMD in family members of controls. Several studies of this nature have suggested that a first-degree relative of an individual with AMD is five times more likely to develop AMD than a family member of a control (7, 8, 9, 10). Twin studies not only confirm this genetic association but also demonstrate striking phenotypic similarities between monozygotic twins, which intuitively demonstrate the major role genes play in this condition (Fig. 31-1) (11).
INITIAL GENETIC STUDIES OF AGE-RELATED MACULAR DEGENERATION
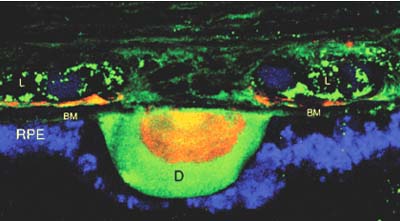
Candidate gene studies (12) have been successful for complex diseases but rely on having the correct identity of the gene or pathway as a biologic basis for that condition predicted in advance. For the study of AMD, candidate gene studies were initially performed on genes implicated in Mendelian maculopathies, such as Best disease (13), Pattern dystrophy (14), Sorsby fundus dystrophy (14), Malattia Leventinese (15), and Stargardt disease (16–19). These diseases were chosen because they recapitulate many features of AMD, such as the deposition of drusenoid-like material, atrophy or choroidal neovascularization (CNV) (Fig. 31-2). To date, these genes have been associated with only a small fraction of the total cases of AMD (2%-3% or less) or have been inconclusive (16, 20).
Linkage mapping studies have the advantage of being hypothesis free and have enjoyed great success in the study of Mendelian conditions but have suffered when applied to complex disorders like AMD due to a lack of statistical power to detect mild effects and lack of resolution produced by low marker density. As an example, the PPAR gene association with diabetes (21) would never have been identified with a linkage approach as it would have required the sample population to include over one million affected sibling pairs to generate this association. Linkage studies for AMD have found only suggestive areas of linkage by standard statistical criteria (22, 23). However, both the CFH gene and LOC387715/HTRA1 locus were found within previously mapped linkage loci (1q31 and 10q26, respectively).
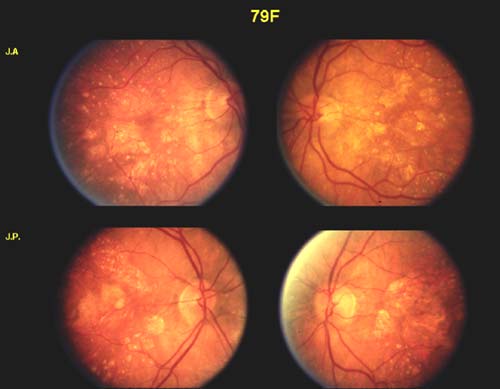
Figure 31-1. Fundus photographs from monozygotic twins with age-related macular degeneration. (From Klein ML, Mauldin WM, Stoumbos VD. Heredity and age-related macular degeneration. Observations in monozygotic twins. Arch Ophthalmol 1994;112:932–937.)
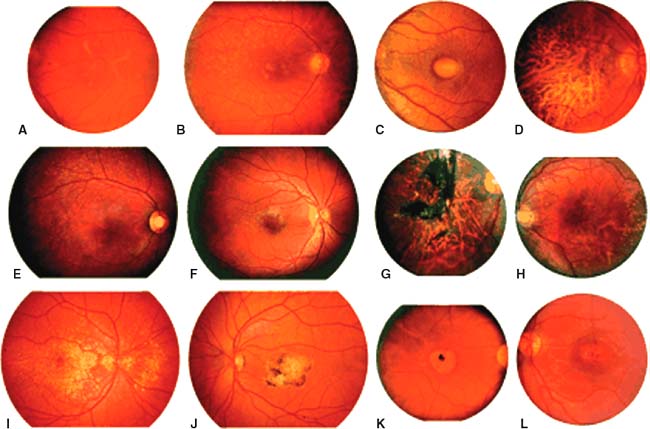
Figure 31-2. Ophthalmoscopic phenotypes of patients affected with molecularly confirmed early-onset heritable macular dystrophies. The left panel of each pair shows the more typical clinical appearance of each disease while the right panel depicts a variant appearance that could be confused with AMD or one of the other heritable dystrophies. A and B: RDS-associated pattern dystrophy (RDS Gly167Asp); C and D: Best disease (VMD2 Ala243Thr); E: Stargardt disease (ABCA4 Phe608Ile/Leu2027Phe); F: Stargardt disease (ABCA4 Cys1488Phe/Cys2150Tyr); G: Sorsby fundus dystrophy (TIMP3 Gly167Cys; photograph courtesy of Dr Samuel Jacobson); H: Sorsby fundus dystrophy (TIMP3 Ser181Cys; photograph courtesy of Dr Michael Klein); I and J: ML (EFEMP1 Arg345Trp); K and L: Stargardt-like dominant macular dystrophy (ELOVL4 common 5 bp deletion). (From Stone EM, Sheffield VC, Hageman GS. Molecular genetics of age-related macular degeneration. Hum Mol Genet 2001;10:2285–2292.)
GENOME-WIDE ASSOCIATION STUDIES
Such challenges have spurred recent interest in the application of GWA studies using single nucleotide polymorphisms (SNPs) as an approach that has a better capability to localize disease susceptibility genes harboring smaller effects. Two GWA studies for AMD using drusen and neovascularization as the primary phenotypes have implicated two major genes in this disease. The first GWA study discovered variants in the CFH gene (3), and the second GWA study discovered variants in the LOC387715/HTRA1 locus (4). Fig. 31-3 shows results from the GWA study indicating association with the CFH gene. Subsequent resequencing based on the principle of linkage disequilibrium (LD) showed that the causal mutation in CFH may be a missense variant in amino acid residue 402 where a tyrosine is replaced by a histidine.
When one looks at the successful design of GWA studies, one immediately appreciates that cases and controls were ascertained from clusters of subjects at the two extremes (Fig. 31-4). In the Klein study (11), cases were comprised of subjects with end-stage AMD as defined by the presence of geographic atrophy or CNV with controls having little to no drusen, which at a mean age of 80 turns out to be typically more difficult to ascertain than AMD patients. The DeWan study (4) was designed to ascertain only Asian individuals given their higher incidence of polypoidal CNV in the absence of drusen. These individuals were compared to age matched controls with no ophthalmoscopic findings of AMD. This thoughtful design enabled the identification of a novel association in the LOC387715/HTRA1 locus, which was not discerned in prior analyses due to greater “noise.”
In summary, to achieve success in GWA studies, study power needs to be increased. There are two ways to do this. The first is by increasing the subject number, as in the WTCC study (24), and future genetic studies of AMD will likely involve tens of thousands of subjects. Secondly, and possibly more importantly, clinical insight plays a critical role in the study design because well characterized, homogeneous cohorts at polar ends of the disease spectrum also increase statistical power. The bottom line with genetic association studies is that, from a phenotypic standpoint, if you put “garbage in” you will get “garbage out.” An example is the lack of associations to date for heterogeneous phenotypic conditions such as primary open-angle glaucoma, for which this approach has not yet been successful. In contradistinction, GWA studies have successfully identified multiple SNPs in the 15q24.1 region as an important risk factor for pseudoexfoliation glaucoma (25), which is a much cleaner, more homogenous phenotype.
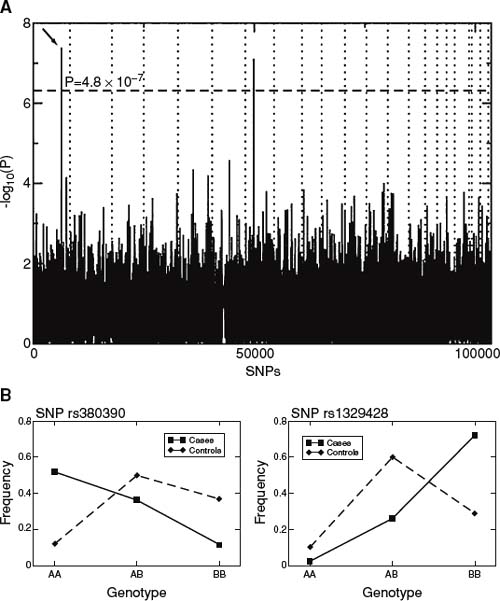
Figure 31-3. A: P values of genome-wide association scan for genes that affect the risk of developing AMD. -log10(p) is plotted for each SNP in chromosomal order. The spacing between SNPs on the plot is uniform and does not reflect distances between SNPs on the chromosomes. The dotted horizontal line shows the cutoff for p ∇ .05 after Bonferroni correction. The vertical dotted lines show chromosomal boundaries. The arrow indicates the peak for SNP rs380390, the most significant association, which was studied further. B: Variation in genotype frequencies between cases and controls. (From Klein RJ, Zeiss C, Chew EY, et al. Complement Factor H Polymorphism in Age-Related Macular Degeneration. Science 2005;308:385–389.)
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


