General Principles of Immunology
Peter Benincasa
Richard Luka
Leonard Bielory
The immune response evolved to protect multicellular organisms from pathogens that range from viruses, bacteria, fungi and protozoa. For each of the various infectious pathogens the immune system has developed characteristic responses that overlap (e.g., predominantly T-cell response for viral infections; a B-cell response with IgM and IgG antibodies for bacterial infections; a macrophage with mixed T and B cells for fungal infections; and an eosinophil, IgE and mast cell response to parasitic infections.1,2 However, a broader view would be to define immunity as a host system’s response to molecules identified as foreign without implying either positive or negative physiologic consequences. This concept would then encompass immune regulation with “appropriate” responses resulting in beneficial outcomes for the host (i.e., control of infection, tumor surveillance) and “inappropriate” responses resulting in deleterious outcomes (i.e., autoimmune disease, immediate hypersensitivity).3 The most common clinically visible effect of the immune response is the development of itis, inflammation that is composed of cardinal features of rubor (erythema), calor (heat), dolor (pain), and tumor (swelling) leading to function laesa (loss of function).
There are four cardinal features of the immunity: (a) the specific recognition of molecules capable of evoking an immune response, (b) the capacity for memory, (c) the subsequent ability to amplify the response, and (d) the ability to discriminate self from nonself. Immune recognition will lead to either immunity or tolerance (i.e., specific unresponsiveness), which is crucial in preventing uncontrolled autoimmune disease.1,2
This chapter will introduce current concepts in basic immunology and provide a framework for understanding immune-mediated disease and the clinical applications of immunomodulation. For the sake of clarity, we will discuss many of the components of the immune system in isolation; however, we should remember that immune regulation is a balance of a myriad of cellular interactions, feedback loops, and molecular cascades.
Innate versus Adaptive Immunity
The immune response has classically been divided into innate and adaptive (or acquired) immunity. Innate immunity is present before exposure to foreign molecules, such as microorganisms, and represents a first line of defense for the individual as its elements act to eliminate the microbe and stimulate a more specific and amplified adaptive immune response.4,5 The chief components of innate immunity include various cellular and molecular components such as the mononuclear phagocyte system including neutrophils, macrophages, mast cells and NK cells, cytokines, complement proteins, physiochemical barriers and both soluble and cell-surface receptors that bind highly conserved molecular patterns present on invading pathogens.28
There are a variety of soluble factors that are not specifically targeted towards any single infectious agent, but have antiinfective properties. These include the protein lysozyme, the interferon proteins and complements of the complement system. Lysozyme is a hydrolytic enzyme found in mucus secretions including tear fluid and is able to cleave the peptidoglycan layer of bacterial cell walls. Interferons comprise a group of proteins produced by virally infected cells that bind to nearly all cells and induce a generalized antiviral state. Complement are a group of serum proteins that circulate in a relatively inactive state, but are converted into active states by various specific (antibody mediated) and nonspecific (disturbed cell membranes) immunologic mechanisms with the ability to damage the membranes of pathogenic organisms. Thus complement bridges the innate and adaptive immune systems. Some of the innate molecules may not be free floating, but associated with immune cell membranes such as Toll-like receptors (see the subsequent text).
Adaptive immunity is defined by the specific recognition of foreign molecules by lymphocytes and their products. Before exposure to antigens, antigen-specific lymphocytes develop that are capable of recognition and response to foreign molecules. This represents the primary response. Activation of lymphocytes results in immunologic memory with the resultant capacity for dramatic amplification of specific and nonspecific effector function on reexposure to the offending molecules, more commonly known as the secondary response. The immune system is capable of recognizing more than 109 antigenic determinants. This remarkably large repertoire results from variability at the antigen-binding sites of immunoglobulins and T-cell receptors (TCRs).1
Adaptive immunity can be further subdivided into humoral and cell-mediated immunity. Humoral immunity is primarily mediated by immunoglobulins, the products of B lymphocytes, either in secreted form or as membrane-bound cell surface receptors. T lymphocytes are primarily responsible for cell-mediated immunity. Recognize, however, that the interaction between B and T cells is crucial in both forms of immunity. Distinctive features of innate verses adaptive immunity are shown in Table 1.6
TABLE 1 | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
An adaptive immune response typically follows exposure to a foreign antigen, conferring memory and the capacity for a secondary response. This has been termed active immunity. Passive immunity describes the transfer of pooled immunoglobulin from immunized individuals to immediately confer immunity without waiting the typical 1 to 4 weeks for the development of an active response. Passive immunity rapidly wanes and does not produce memory.1
Overall, coordination of the innate and adaptive immune responses is crucial for the vitality of the body and the individual organs including the eye.
Toll-Like Receptors
The innate immune response maintains a capacity to distinguish pathogens from self proteins. A variety of soluble and cell-surface receptors bind highly conserved pathogen-associated molecular patterns found on microorganisms. Mannose-binding lectins and ficolins are circulating effector proteins that bind mannose structures and N-acetyl glucosamine on microorganisms eventually leading to activation of the classical pathway of the complement system. Toll-like receptors (TLRs) are cell-surface and intracellular pattern recognition receptors discovered in the 1990s and so named because of their homology to the Toll genes of Drosophila. TLRs are expressed on a variety of different cell types, which first come into contact with pathogens from the outer environment including dendritic cells,7 macrophages, endothelial and epithelial cells as well as other effector cells.8,9 Ten TLRs have been identified in humans and are characterized according to their agonists, either bacterial products, fungal products or nucleic acids.10 These TLR ligands are widely expressed on microbial but not mammalian cells. TLRs which recognize bacterial products tend to be expressed on the cell surface, while those recognizing nucleic acids reside on endosomal membranes.10 The binding of a TLR to its ligand leads to intracellular signaling via an adaptor molecule most commonly MyD88 and then to the activation of NF-κB with the resultant expression of pro-inflammatory and regulatory cytokines such as IFN-γ, TNF-α, IL-1β, and IL-12. This cytokine release then leads to the recruitment and activation of a variety of inflammatory cells interfacing and instructing the adaptive immune response in infectious and allergic ocular disorders.11,12,13,31
There is widespread TLR expression in the eye.13 TLRs on the corneal and conjunctival epithelium and stroma have been demonstrated to participate in a variety of pathologic processes. TLR-4-expressing corneal fibroblasts have been shown to play a role in lipopolysaccharide-induced inflammation in corneal ulcers. In vernal keratoconjunctivitis there is modification of the normal expression of TLRs with TLR-4 being significantly upregulated and TLR-9 significantly downregulated though the mechanism is unclear. TLR-9 agonists (CPG oligonucleotides) have been shown to bias the immune response toward a TH1 response and away from the TH2 response seen in conjunctival allergy and VKC.12,13,32
Antigen
Antigens are molecules capable of specifically binding to lymphocyte receptors. More precisely, immunogens are those antigens whose binding evokes an immune response (although the term antigen still is commonly used in its place). The term allergen is commonly used for those specific immunogens (antigens) that bind specifically to IgE antibodies lead to the “allergic rsponse. Whereas immunoglobulins are capable of binding soluble antigens, TCRs bind only processed peptides presented in the context of specific cell surface proteins, collectively called the major histocompatibility complex (MHC) gene products. The specific site on an antigen that is bound by an antibody is termed a determinant or epitope. Haptens are simple molecules, commonly less than eight amino acids in size that can bind to antibodies, but cannot evoke an immune response unless bound to a macromolecule. In this instance, the hapten behaves like an exogenous determinant on the larger carrier protein.1,2,14
The typical immune response is downregulated by a number of mechanisms and so wanes with time. The success of the immune response in eliminating its antigenic stimulus, the natural end-differentiation of lymphocytes into memory cells or short-lived cells, and feedback inhibition in the form of cytokine regulatory cascades and perhaps the development of anti-idiotypic antibodies all play a role in this self-regulation. Tolerance, or learned immunologic unresponsiveness, is discussed later.
Components of the Immune System
The cells and lymphoid organs that compose the immune system participate in a series of complex interactions to regulate the immune response (Fig. 1). In this section, we outline the essential components and their functions before a more explicit discussion of their role in immune-mediated disease later in this chapter. The cellular components of the immune system of the eye have many similarities with the systemic immune system as reflected by the conjunctiva associated lymphoid tissue (CALT) which is histologically very similar to the gut associated lymphoid tissue or the bronchial associated lymphoid tissue.34
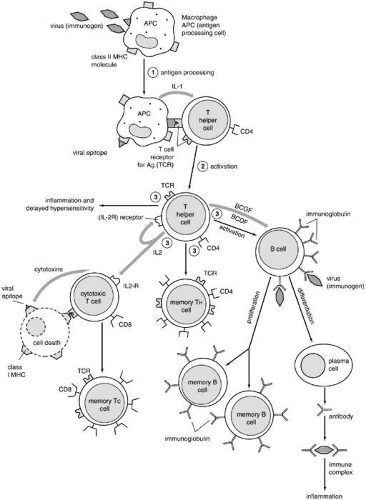 FIGURE 1. Grand scheme of adaptive immune system. (From Goodman JW. The immune response. In Stites DP, Terr AI (eds). Basic and Clinical Immunology, 7th ed. Norwalk, CT: Appleton and Lange, 1991:36 with permission.) |
Physiochemical Barriers
Physiochemical barriers are significant first-line elements of a host’s defense. The skin and mucous membranes limit access to the host by invading microorganisms. Enzymes, such as lysozyme, nonspecifically inhibit microbial growth. In the eye, the conjunctiva and the tear fluid layer provide the primary barrier against environmental aeroallergens, chemicals, and infectious agents. The tear fluid layer contains both specific and nonspecific immunologically active proteins, including lysozyme, interferons, histamine, tryptase, lactoferrin, ceruloplasmin, vitronectin, immunoglobulin A (IgA), immunoglobulin G (IgG), immunoglobulin M (IgM), and immunoglobulin E (IgE).4
Lymphocytes
Lymphocytes are the cells responsible for the specificity of immune recognition and for coordination of the immune response. They are derived from a lymphoid progenitor cell in the bone marrow and are divided into three classes: T cells (lymphocytes), B cells (lymphocytes), and natural killer (NK) cells. T cells and B cells are small 8- to 10-μm-in-diameter lymphocytes with large nuclei that are morphologically indistinguishable, but functionally distinct that can easily be differentiated by the cell surface proteins they express: T cells with CD3, CD4, and CD8 and B cells with CD-19 surface markers.1
NK cells are large lymphocytes with many cytoplasmic granules, distinct cell surface markers (CD16), and the ability to lyse cells directly, especially tumor cells or normal cells infected by virus. NK cells lack immunoglobulin or TCRs for antigen recognition yet do not kill their targets at random. Their means for target cell recognition is not well understood. NK cells do participate in antibody-dependent cellular cytotoxicity (ADCC) on the basis of their expression of a low-affinity receptor for the Fc portion of IgG (CD16). In ADCC, a target cell coated with IgG can be lysed directly by an NK cell after binding of the Fc portion of IgG with its low-affinity receptor CD16. This not only provides a means of recognition for the target by the NK cell, but also serves to activate it to release its granule contents such as perforin, an enzyme that perforates the wall of the target cell, and secrete cytokines to augment the inflammatory response.1
B cells develop from their precursors in the fetal liver and adult bone marrow. (They were first shown to mature in birds in the bursa of Fabricius, hence “B” cell.) They are primarily responsible for humoral immunity and are the exclusive producers of immunoglobulins, also known as antibodies, thus playing a vital role in the recognition and elimination of foreign antigen (further discussion on immunoglobulins below). Mature B cells can be divided into memory cells for the development of a rapid secondary response and plasma cells, which are totally committed to produce a single protein, an immunoglobulin. Plasma cells are terminally differentiated producers of large amounts of antibody (Fig. 2). B cells also interact closely with helper T cells through cell surface proteins such as CD40 and class II MHC and their complementary ligands.
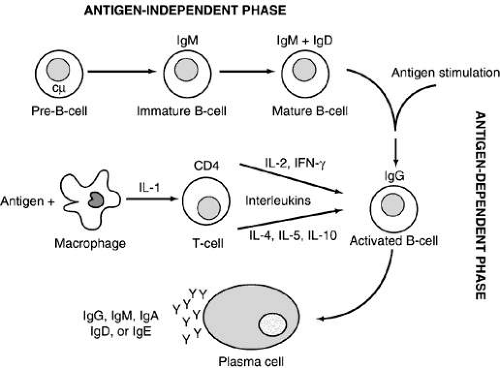 FIGURE 2. B-cell differentiation pathway. Pre-B cells characterized by intracytoplasmic immunoglobulin M (IgM) heavy chain (cμ) differentiate into immature B cells. These B cells are the first cells to reproduce light chains, which can then combine with the μ heavy chain to become expressed as IgM on the cell surface. The immature B cell differentiates into a mature B cell, which expresses both IgM and immunoglobulin D (IgD) on its cell surface. This part of the differentiation process does not require antigen and is referred to as antigen independent. On contact with the specific antigen via the surface immunoglobulin, the mature B cell becomes activated. Together with helper T cells (CD4+) and interleukins/cytokines derived from T cells and monocytes, the activated B cell matures into a plasma cell to produce immunoglobulins of a single isotype (Y). However, an antigen stimulates a polyclonal B-cell response, which results in many plasma cells and immunoglobulin production of several classes or isotypes. (From Medical Knowledge Self-Assessment Program [MKSAP]: Allergy and Immunology. Philadelphia: American College of Physicians, 1993:155 with permission.) |
T cells develop from their bone marrow precursors in the thymus, where somatic gene rearrangement gives rise to functional TCR complexes as well as distinctive cell surface proteins. T cells can be functionally subdivided into either helper or cytolytic (CTL) cells. These functionally different populations express distinct cell surface proteins: CD4 on helper cells and CD8 on CTLs, which serve as ligands for the MHC gene products on antigen-presenting cells.5
CD4+ cells play a vital role in B-cell growth and differentiation, including the production of antibodies.6 They also are crucial in macrophage activation and in upregulating or downregulating the immune response. CD4+ cells can be further subdivided into TH1 and TH2 lymphocyte populations based on their cytokine-producing properties. Cytokines are small proteins produced by cells in response to a variety of inducing stimuli. Cytokines are produced by their producer cells and then influence the behavior on target cells. Hormones are classical polypeptides that also fulfill this definition, but are not by convention classified as cytokines since they are produced by specific endocrine organs (e.g., the thyroid gland), whereas a cytokine may be produced by more than one cell type in a number of different tissues. Cytokines similar to hormones act on the cells that produce them (autocrine), on cells in a distant organ (endocrine), or on cells in the immediate vicinity of their production (paracrine) fashion to regulate lymphocytes, antigen-presenting cells, other inflammatory cells, and immunologically active molecules involved in cellular communication. Multiple studies have shown that cytokines also are characterized by their ability to act on different cells (pleiotropism), produce different effects on the same target cell, share properties with other cytokines (redundancy), and influence the production of other cytokines.1 It also has been discovered that among their many functions, cytokines can affect cell growth and differentiation similarly to those of the growth factors. Given the enormous diversity of cells producing cytokines and the numerous effects (many of which still are unknown) exerted by these molecules, it has been difficult to develop a satisfactory classification (Table 2). Cytokines also may be classified by cell of origin, but it is now clear that certain cytokines are produced by many cells, making these criteria impractical for classification.
TABLE 2. Cytokine Families | ||
|---|---|---|
|
There are preferential patterns of T-lymphocyte production of cytokines that generate an immunological dimorphism between Th1 and Th2 activating the most efficient way to eliminate a specific pathogen.14 TH1 lymphocytes express inflammatory cytokines that are involved in the effector functions of cell-mediated immunity with the primary production of cytokines that include interferon-γ (IFN-γ) and interleukin (IL)-2. TH2 lymphocytes produce cytokines such as IL-4 and IL-10, which often antagonize the inflammatory effects of IFN-γ and stimulate B-cell differentiation. IL-4 is required for the production of antibodies, IgA, and IgE, primarily associated with mucosal surfaces as this cytokine stimulates isotype switching to the α and ε heavy chains. TH1 lymphocytes appear to be more important in the defense against intracellular pathogens, whereas TH2 lymphocytes have an important role in the protection against parasitic disease. These two cell populations also are responsible for different pathologic states with TH1 responses involved in organ-specific autoimmune disorders, including experimental autoimmune uveoretinitis, and TH2 cells involved in atopic disease.14 Indeed in the heterogeneous group of allergic ocular diseases that involve a variety of different pathogenetic mechanisms, cytokines and cell populations the relative role of TH1 and TH2 pathways has been defined. Newer techniques are being developed to more accurately assess inflammatory mechanisms involved in ocular diseases that may aid in diagnosis and treatment.35
CD8+ lymphocytes are primarily responsible for cytolysis of virus-infected cells, malignant cells, and tissue allografts. This type of cell killing is highly specific as antigenic peptides are presented to the CD8+ cell in the context of class I MHC molecules and require direct contact of the CTL with the target cell. CD8+ cells also participate in cell-mediated immunity by activating macrophages. The role of CD8+ cells as “suppressor” cells is less clear, although a distinct subpopulation of non-CTL CD8+ cells may be distinguished.
Mononuclear Phagocytes
Cells of the mononuclear phagocyte system are derived from a common myeloid progenitor cell in the bone marrow. Their primary functions are phagocytosis and antigen presentation. In the peripheral blood, they are found as incompletely differentiated cells known as monocytes. Once in tissue, they mature and are known as macrophages.
Macrophages function as important effector cells of innate immunity. They are capable of phagocytizing foreign particles and microorganisms and killing them with lysosomal enzymes. They also produce and secrete cytokines that recruit other inflammatory cells and promote the inflammatory response. Among the cytokines typically expressed by macrophages are tumor necrosis factor (TNF)-α, IFN-γ, IL-1, and IL-6.
Macrophages also act as efficient antigen-presenting and costimulatory cells for T cells. T cells, in turn, may secrete cytokines, especially IFN-γ, which activate macrophages, upregulating their killing ability and antigen-presenting capacity.
Dendritic Cells
Dendritic cells are extremely efficient antigen-presenting cells with distinctive spinelike projections. Interdigitating dendritic cells are bone marrow-derived and are found in the interstitium of most organs. In the skin and conjunctiva, they are known as Langerhans cells and bear the CD1 cell surface marker. Langerhans cells are extremely mobile, migrating from the skin to the peripheral lymph nodes, and are particularly effective at presenting antigen to CD4+ cells. Interaction between antigen, dendritic cell and toll-like receptors participate in the preferential differentiation of T cells into Th1 or Th2 effector cells (Fig. 3).
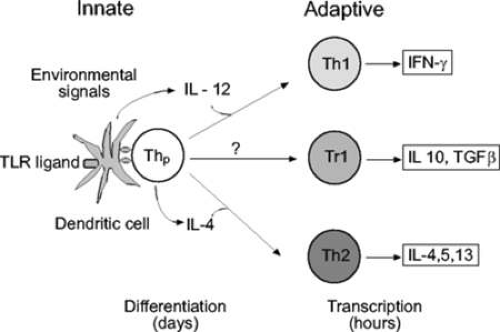 FIGURE 3. Regulation of T-cell differentiation by signals provided by dendritic cells (DCs). Strong toll-like receptor (TLR) signaling enhances IL-12 production leading to T-helper cell (Th) type-1 differentiation. Th2 differentiation, in contrast, results in the absence of IL-12 when IL-4 gene transcription in naïve T-cells is promoted. Whether there are DC-derived signals that can actively promote Th2 differentiation is not entirely clear. Naïve T-cells can also differentiate in the periphery into regulatory phenotypes that can down regulate immunity: the signals involved in regulatory T-cell differentiation are not known but may include IL-10 (not shown). IFN: interferon; TGF-β: transforming growth factor-β. (Georas, S. N., Guo J., De Fanis U., and Casolaro V. 2005). |
Follicular dendritic cells are found in the germinal centers of lymph nodes, the spleen, and the mucosal lymphoid tissue. They are unrelated to interdigitating dendritic cells, but also are extremely important antigen-presenting cells within lymphoid tissue.1
Neutrophils
Leukocytes containing abundant cytoplasmic granules play vital roles in the elimination of microorganisms and in the acute inflammatory response. Neutrophils comprise more than 90% of granulocytes and between 60% and 70% of the circulating leukocyte pool. They are extremely mobile, responding rapidly to chemotactic stimuli, adhering to specific ligands on the endothelial lining of blood vessels, and, via diapedesis, extravasating into tissue sites of injury. Chemoattractants for neutrophils include the byproducts of bacterial metabolism; C5a, which is a byproduct protein component of the complement cascade system; as well as leukotriene B4 and platelet- activating factor (PAF), both derived from cell membrane lipids.8
Various cellular processes including adhesion, migration, proliferation, differentiation, and activation are modulated by cell adhesion molecules (CAMs). Adhesion molecules are expressed on leukocytes, vascular endothelium, and epithelial cells and have been identified within various structures of the eye including the cornea, conjunctiva, choroid, uvea, and optic nerve.15,9 They are grouped in four major families: the integrins, the selectins, the cadherins, and the immunoglobulin supergene family.16,17,18 Endothelial CAMs are important in regulating the infiltration of neutrophils into sites of inflammation include E-selectin and intercellular adhesion molecule-1 (ICAM-1). ICAM-1 binds to a complement receptor known as membrane attack complex type 1 (MAC-1) (i.e., complement receptor 3 or CR3) on the neutrophil, establishing firm adhesion and leading to extravasation.19 ICAM-1 also plays a role in ocular hypersensitivity reactions as reflected by its increased expression in atopic patients during allergen-specific conjunctival challenge15,19,20,21,22,23.12
During phagocytosis, neutrophils engulf invading microorganisms, isolating them in phagosomes. Cytoplasmic granules containing a variety of cytotoxic enzymes fuse with the phagosome and destroy the pathogen. Most invading microorganisms are coated, or opsonized, with IgG or another complement protein byproduct known as iC3b. These can then bind to specific receptors on neutrophils (FcRII and MAC-1), facilitating phagocytosis.
The neutrophil has both oxygen-dependent and oxygen-independent antimicrobial systems. The oxygen-dependent system produces reactive oxygen metabolites (“respiratory burst”) that are toxic to microorganisms. The oxygen-independent system relies on the cationic peptides and enzymes released from cytoplasmic granules for killing.
Eosinophils
Eosinophils are important effector cells in atopic disease and parasitic infection24,25 They contain a bilobed nucleus and distinctive acidophilic granules and are far more prevalent in tissue than in peripheral blood. Their cytoplasmic granules contain a number of basic proteins, including major basic protein (MBP), eosinophil cationic protein (ECP), eosinophil-derived neurotoxin (EDN), and eosinophil peroxidase (EPO), that are responsible for the eosinophils uptake of acid stains. MBP makes up more than 50% of total granule protein, is known to be extremely toxic to parasites, and is a vital component of eosinophil ADCC. MBP also is responsible for damage to epithelial cells, as seen in the respiratory epithelium in patients with asthma, and which is similar to the damage in the corneal epithelium in patients with vernal and atopic keratoconjunctivitis.
IL-5, produced by the TH2 subset of CD4+ cells and activated mast cells, is a potent stimulant for eosinophil production and activation. It also prolongs eosinophil survival and causes secretion of granule proteins. IL-4, another TH2 cytokine important in immediate hypersensitivity disease, causes endothelial cells to upregulate the expression of the adhesion molecule vascular cellular adhesion molecule (VCAM-1), which binds to its ligand VLA-4 on eosinophils. This results in eosinophil recruitment to sites of immediate and chronic immunologic hypersensitivity inflammatory reactions.
Mast Cells and Basophils
Mast cells and basophils play important roles in mediating immediate hypersensitivity. They each express large numbers of high-affinity IgE receptors (FcεRI) on their cell surfaces. Mast cells are localized to mucosa, epithelial surfaces, and connective tissue. Basophils may be considered the peripheral blood counterpart to the mast cell.25
Mast cells can be divided into two types based on their expression of secretory granules: tryptase mast cells (MCT) and tryptase-chymase mast cells (MCTC). MCT are found primarily in the lung and intestinal mucosa, whereas MCTC are localized to the skin and intestinal submucosa. The predominant form of mast cells found in the normal conjunctiva is MCTC; however, there is a noticeable increase in the MCT type in chronic conjunctival inflammatory conditions. It is estimated that more than 50 million mast cells are present in the conjunctiva.
Mast cells and basophils are activated by the crosslinking of FcεRI molecules on their surface after the binding of multivalent antigen to sufficient IgE. Activated mast cells release their preformed mediators in a regulated fashion and then synthesize lipid-derived mediators of inflammation.
Stem cell factor (SCF) is an essential growth factor for mast cells and enhances IgE-dependent mast cell mediator release in addition to the various cytokines that are generated and released. The mast cell stores and manufactures SCF and in mast cell mediated disorders like seasonal allergic conjunctivitis levels have been found to be increased.
The prototype preformed vasoactive mediator released by mast cells is histamine. Histamine binding to its cell receptors results in vasodilation, endothelial cell contraction, and associated plasma leakage, as well as bronchial and intestinal smooth muscle constriction.26. Four major subtypes of histamine receptors, H(1) to H(4), have been identified and appear to differ in their location, second messengers, and histamine-binding characteristics. In addition, different histamine receptor agonists and antagonists appear to bind to different portions of the receptor complex.27 A fifth receptor subtype, the intracellular H(IC), has only been defined by its location within cell types that are not traditionally associated with histamine. Other preformed mediators include tryptase, chymase, carboxypeptidase-A, proteoglycans, eosinophil chemotactic factor, and neutrophil chemotactic factor. Lipid-derived mediators synthesized after activation include leukotrienes B4 (one of the most potent neutrophil chemoattractants known to the human body), C4, D4, and E4 (also known as SRS-A, a slow-releasing substance of anaphylaxis), prostaglandin D2, and PAF, which all have vasodilatory and smooth muscle constrictive effects (Fig. 4).4
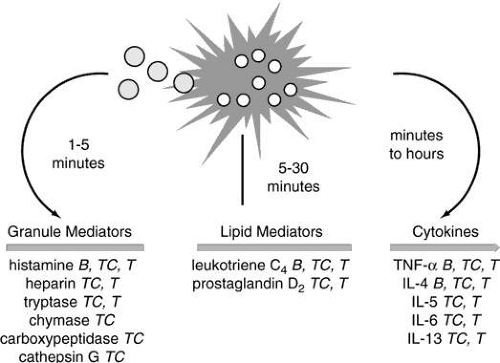 FIGURE 4. Time course of mediator release from human basophils (B), MCTC mast cells (TC), and MCT mast cells (T). Granule mediators are preformed; lipid mediators are newly generated; cytokines can be both preformed and newly generated. (From Medical Knowledge Self-Assessment Program [MKSAP]: Allergy and Immunology, 2nd ed. Philadelphia: American College of Physicians, 1997:23 with permission.) |
Despite their historical and central role in type I hypersensitivity, allergy and acquired immunity, there is increasing evidence and interest in the mast cell’s role in the hosts initial defense against bacterial, viral and fungal antigens. The strategic localization of mast cells in tissues exposed to antigenic assault put them in the ideal position for rapid response. A variety of mechanisms have been described by which the innate immune system can be activated with these cells playing a pivotal role. Complement, immunoglobulin-binding proteins, pathogen associated mannose-binding proteins and most recently toll-like receptors all share in determining the activation of innate immunity through mast cells.28,29,30,31
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


