Chapter 2 Clinical embryology and development of the eye
The vertebrate eye is formed through coordinated interactions between neuroepithelium, surface ectoderm, and extraocular mesenchyme.1 The neuroectoderm gives rise to the retina, iris, and optic nerve; the surface ectoderm forms the lens and corneal epithelium; the extraocular mesenchyme comprising mesodermal and neural crest cells gives rise to the corneal stroma, corneal endothelium, extraocular muscles, and fibrous and vascular coats of the eye.2
Three main periods can be distinguished in the prenatal development of the eye:
1. Embryogenesis includes the establishment of the primary organ rudiments and finishes when the optic groove (optic sulcus), which is considered the anlage of the eye, appears on either side of the midline at the expanded cranial end of the open neural folds around the end of the third gestational week.
2. Organogenesis includes the development of the primary organ rudiments and extends to the end of the eighth week.
3. Differentiation involves the differentiation of the primitive organs into a fully or partially active organ starting at the beginning of the third month. During this period, the retina, optic nerve, and anterior rim of the optic cup mature and the vitreous, lens, and angle structures develop3 (Table 2.1, see also Chapter 4 and 6).
Table 2.1 Overview of eye development4
| Gestational age | Developmental milestone |
|---|---|
| 22 days | Optic primordia appears |
| 2nd month | Hyaloid artery fills embryonic fissure |
| Closure of embryonic fissure begins | |
| Lid folds appear | |
| Neural crest cells (corneal endothelium) migrate centrally; corneal stroma follows | |
| Choroidal vasculature starts to develop | |
| Axons from ganglion cells migrate to optic nerve | |
| 3rd month | Sclera condenses |
| Lid folds meet and fuse | |
| 4th month | Retinal vessels grow into nerve fiber layer near optic disc |
| Schlemm’s canal appears | |
| Glands and cilia develop in lids | |
| 5th month | Photoreceptors develop inner segments |
| Lids begin to separate | |
| 6th month | Dilator muscle of iris forms |
| 7th month | Central fovea thins |
| Fibrous lamina cribrosa forms | |
| Choroidal melanocytes produce pigment | |
| 8th month | Iris sphincter develops |
| Chamber angle completes formation | |
| Hyaloid vessels regress | |
| Retinal vessels reach periphery | |
| Myelination of optic nerve fibers is complete to lamina cribrosa | |
| Pupillary membrane disappears |
Eye organogenesis (4th–8th week gestation human)
Fourth week
In the fourth week, the optic pits deepen and form the optic vesicles (OVs) which are evaginations of the lateral walls of the diencephalon. The OVs are connected to the forebrain by the optic stalk (a short tube that eventually forms the optic nerve) (Fig. 2.1A). Interaction between the OV and surface ectoderm (SE) induces the lens placode, and the wall of the OV in contact with the SE thickens to form the retinal disk. Towards the end of the fourth week invagination begins to transform the OV into the optic cup (OC). Simultaneously, the primordia of the extraocular muscles appear as condensations in the periocular mesenchyme. Disruptions in these early steps lead to severe congenital anomalies, including anophthalmia, microphthalmia, and optic fissure closure defects (coloboma).5,6
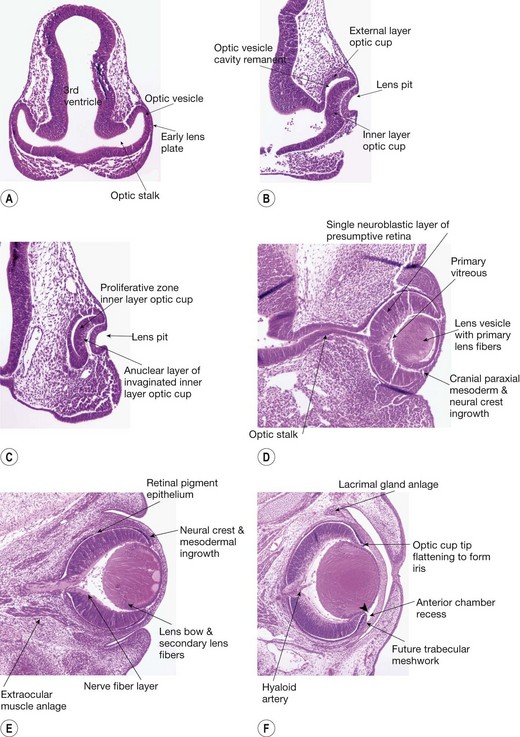
Fifth week
The process of invagination of the OV to form the OC predominates in the fifth week. Invagination involves the retinal disk, lens plate, and the ventrocaudal wall of the OV (Fig. 2.1B). Invagination of the retinal disk of the OV leads to formation of the inner layer of the OC which becomes the neural retina, while the external layer of the OC will become the retinal pigment epithelium (RPE) (Fig. 2.1C). The OC is not continuous, and forms a fold inferiorly and ventrally that is continuous with the optic stalk. This fold, called the embryonic (optic) fissure (Fig. 2.3), allows the passage of the hyaloid artery into the OC. The primary vitreous develops around the hyaloid vasculature. The process of invagination also involves the lens placode (plate), which leads to the formation of the lens pit. The lens pit deepens to become the lens vesicle. Further development leads to the lens vesicle separating from the SE.7 The lens vesicle is large and fills the OC.8 The SE becomes the corneal epithelium (Fig. 2.1D).1,3
Sixth week
In the sixth week the optic fissure closes after the edges of the OC that border the fissure become closely apposed. The pattern of gene expression at those apposed edges must be spatially and temporally appropriate to bring about fusion.9 The embryonic fissure closure begins in the middle and then extends anteriorly and posteriorly (see Fig. 2.3).
The development of the retina progresses with the RPE forming a single layer of cuboidal cells. A primitive Bruch’s membrane arises. The sensory retina thickens due to proliferation of cells in the germinative zone of the inner layer of the OC. At this stage, retinal ganglion cell axons, which form the optic nerve fibers, first enter the optic stalk to exit the primitive eye4 (Fig. 2.1E). The secondary vitreous, a cellular structure with associated extracellular matrix (ECM), forms and remodels the primary vitreous filling the remaining retrolenticular space.10
Seventh week
The main events during the seventh week include the maturation of the RPE, the development of the sensory retina with the formation of outer and inner neuroblastic layers at the posterior pole. Primary lens fibers form to obliterate the cavity in the lens vesicle. The periocular mesenchyme evolves with the formation of the choroidal vasculature posteriorly and the development of the anterior segment (Fig. 2.1F).
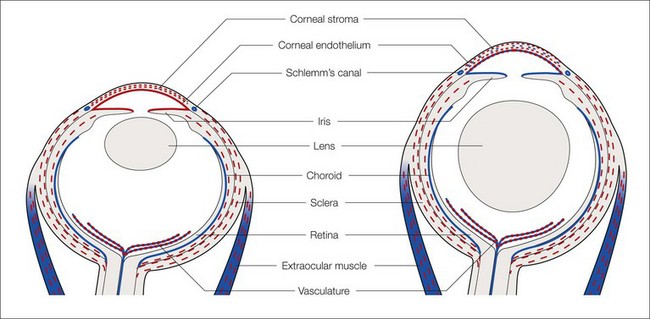
The anterior periocular mesenchyme in the mammal has contributions from neural crest and mesodermal cells, whereas in the chick there are only neural crest contributions. The mesenchymal cells migrate forward so that cells of neural crest and mesodermal origin contribute to the corneal stroma, endothelium, and trabecular meshwork: Schlemm’s canal (SC) is of mesodermal origin (Fig. 2.2).
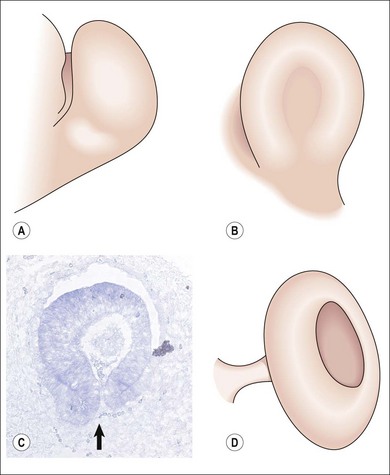
Fig. 2.3 Optic vesicle invagination three dimensional representation (A) Optic vesicle outpouching (9.5 DG mouse 32 DG human), (B) Optic fissure closing (C) Parasaggital section of mouse eye just prior to fissure closure (reproduced from Mihlec et al50) (D) Fusion of optic fissure
(A, B, D reproduced with permission from Fitzpatrick and van Heyningen)5.
Eighth week
In the eighth week, there is marked development of the optic nerve as ganglion cells differentiate; by the end of this week, 2.67 million axons have formed. Optic nerve axons start to make contact with the brain and establish a rudimentary chiasm.11 The RPE nears maturation with the appearance of melanosomes. Müller cells appear now and extend radial fibers inwards to form the internal limiting membrane and outward toward the future external limiting membrane.
The lens develops rapidly during this period. The primary lens fibers fill the lens vesicle. The intracellular organelles disappear. The equatorial epithelial cells begin to divide and new cells are pushed posteriorly, then elongate and become the secondary lens fibers. With the formation of the secondary lens fibers, there is development of the lens bow which represents the nuclei of the secondary lens fibers. They form an arc with a forward convexity (Fig. 2.1E). The lens “sutures” develop where the secondary lens fibers meet in a linear pattern at the anterior and posterior poles of the lens. The sutures initially are in a Y shape anteriorly and an inverted Y ( ) posteriorly.7,12
) posteriorly.7,12
Differentiation and maturation of elements
Cornea
The cornea’s main function as a transparent “window” derives from its unique structure and composition permitting the transmission and refraction of light.13 It consists of three layers: an outer epithelial layer, a middle stromal layer consisting of a collagen-rich ECM interspersed with keratocytes, and an inner endothelial cell layer.
The corneal epithelium develops from SE and its maturation is related to eyelid development. Rudimentary lids fuse 8 weeks after ovulation and do not separate until 26 weeks.14 Bowman’s membrane, which underlies the corneal epithelium, develops from the processes of superficial mesenchymal stromal cells. It first becomes apparent around week 16 and is easily recognizable by the fifth month.15
The beginnings of the corneoscleral junction, which will become the limbus, appear at the end of the eighth week. This is marked by a change in stromal appearance at the periphery of the cornea, with cells acquiring a more polymorphic shape and losing regular orientation. By week 11, this junction is well demarcated. In place of the limbal folds identifiable in adult corneas, a ridge-like structure circumscribes and demarcates the developing cornea. This represents the primitive corneal epithelial stem cell niche.15
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


