, Yasuya Nomura2 and Yasuya Nomura3
(1)
The Society for Promotion of International Oto-Rhino-Laryngology, Tokyo, Japan
(2)
The University of Tokyo, Tokyo, Japan
(3)
Showa University, Tokyo, Japan
Abstract
Blood circulation is of paramount importance to maintaining the functions of the inner ear. However, because of the size and location of the labyrinth in the temporal bone, it is difficult to estimate clinically the details of its circulation. Histochemical staining of alkaline phosphatase allows clear visualization of the capillary networks of the inner ear. Studies of the distribution of capillaries in the inner ear sensory organs have shown that beneath the sensory epithelium dense capillary networks exist in the otolithic organs and semicircular canals. By contrast, the organ of Corti is poorly vascularized. Massive hemorrhage occurs in the inner ear in patients with blood diseases. Temporal bone histopathology reveals that cryoglobulinemia and diabetes mellitus cause pathological changes in the inner ear. A rolled-up tectorial membrane is often seen in viral labyrinthitis, though this finding may not be pathognomonic for viral infection. Vascular disturbances may also cause the formation of an encapsulated mass at the tip of the tectorial membrane, the size of which is generally smaller than those seen in viral labyrinthitis.
Keywords
Alkaline phosphataseBuerger’s diseaseCryoglobulinemiaDiabetes mellitusInner ear blood vesselLeukemiaTakayasu’s arteritis3.1 Anatomy
3.1.1 Arteries and Veins
The artery of the inner ear arises from the vertebral artery system. The labyrinthine artery usually arises from the anterior inferior cerebellar artery (AICA), though occasionally it comes directly from the basilar artery. The endolymphatic sac is supplied with arterial blood from branches of the external carotid artery, i.e., the meningeal branch of the occipital artery and posterior meningeal artery. The latter is a branch of the ascending pharyngeal artery (Fig. 3.1).
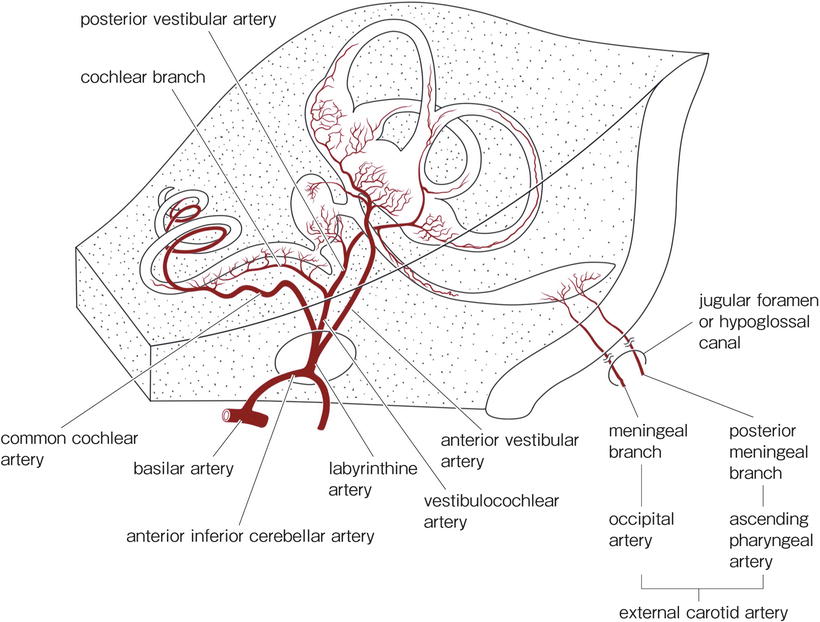
Fig. 3.1
Arteries of the inner ear
The labyrinthine artery divides into the cochlear artery, vestibular artery, and vestibulocochlear artery in the internal auditory canal. Considerable variation exists in the exact manner of branching and distribution of the labyrinthine artery and its tributaries. The diameter of the labyrinthine artery is approximately 0.3 mm in humans.
The veins of the inner ear do not run through the internal auditory canal, with rare exceptions (Fig. 3.2). The vein of the cochlear aqueduct (inferior cochlear vein) runs within the canal of Cotugno and drains most of the venous blood from the cochlea into the inferior petrosal sinus or the superior bulb of the internal jugular vein. The vein of the vestibular aqueduct runs within the paravestibular canaliculus to join the sigmoid sinus. The venous drainage of the vestibular labyrinth is via the vein of the cochlear aqueduct and the vein of the vestibular aqueduct.
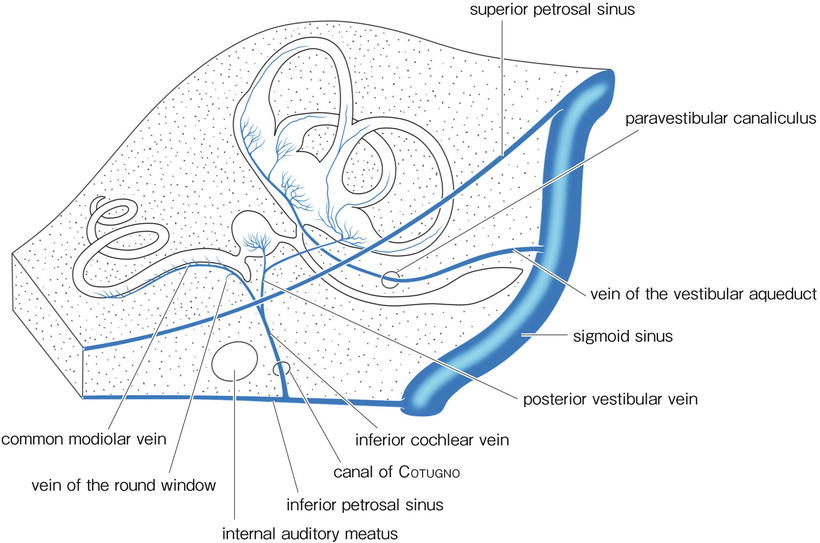
Fig. 3.2
Veins of the inner ear
In the modiolus, modiolar arteries become tortuous before reaching the membranous labyrinth and the spiral ganglion (coiled arteriole). The interscalar septum separates the scala vestibuli from the scala tympani of the adjacent upper turn. It conveys radiating arterioles to the spiral ligament and stria vascularis. Venules do not accompany the arterioles, but run in spiral form in the septum (Figs. 3.3 and 3.6).
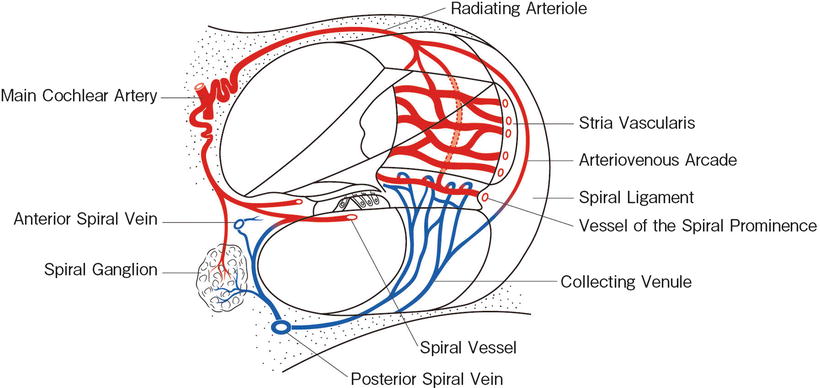
Fig. 3.3
Vascular system of the cochlea
In the interscalar septum, small bony canals radiate from the modiolus laterally to the spiral ligament, forming networks throughout the septum (Figs. 3.4 and 3.5). Arterioles and capillaries run within these canals. These vessels may be observed in a surface specimen of the interscalar septum after decalcification, dissection, and staining [1].
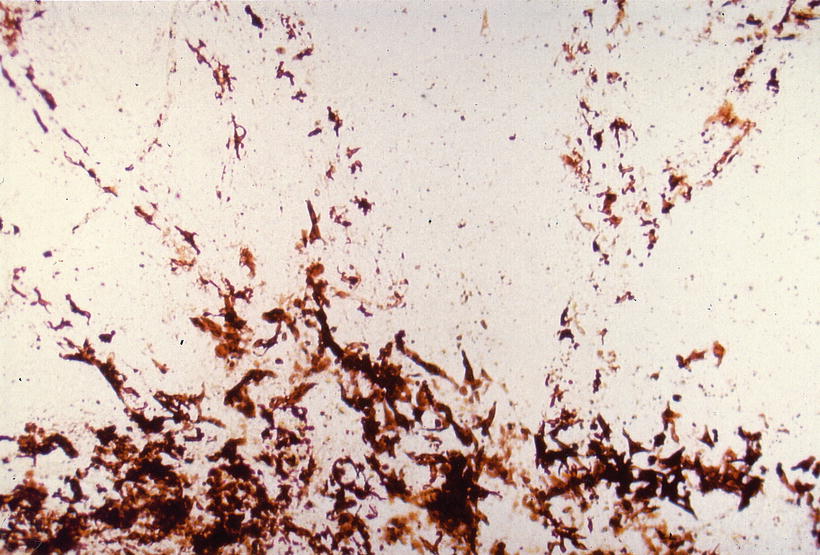
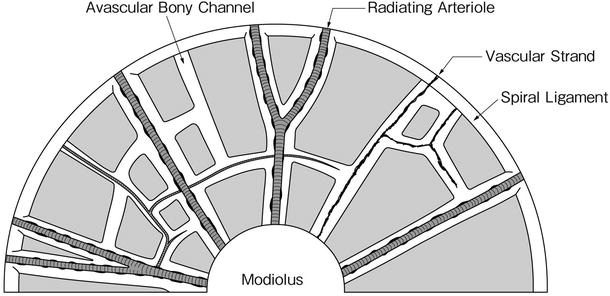
Fig. 3.4
Pigment cells in the interscalar septum, indicating the existence of canals. Abundant pigment cells are found in the modiolus (below). Fifty-year-old man, surface preparation, no stain (original ×6.5) [1]
Fig. 3.5
Schema of the canalicular system in the human interscalar septum
Arterioles run through the center of the wide bony canal. In other words, arterioles have a wide perivascular space. Pigmented cells are often seen attached to the arterioles. The cells contain melanosomes and premelanosomes, although the functional roles of the pigmented cells are not clear (Figs. 3.6, 3.7, 3.8, and 3.9).
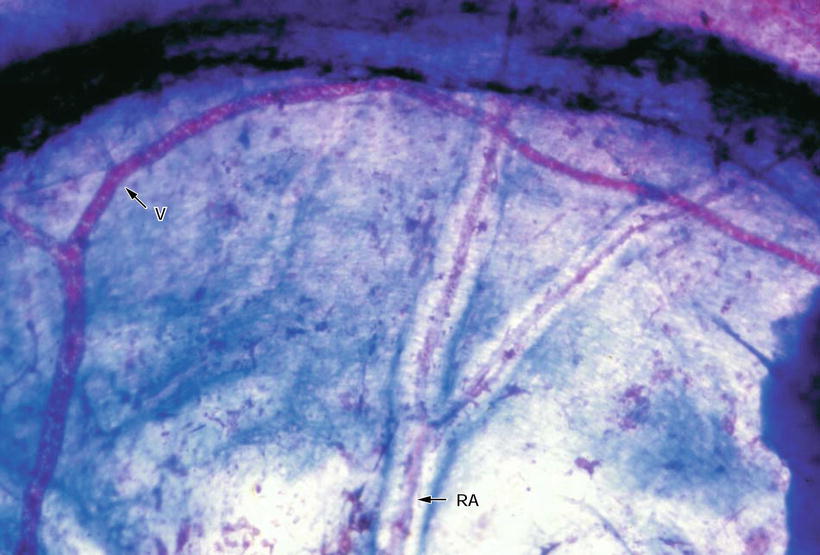
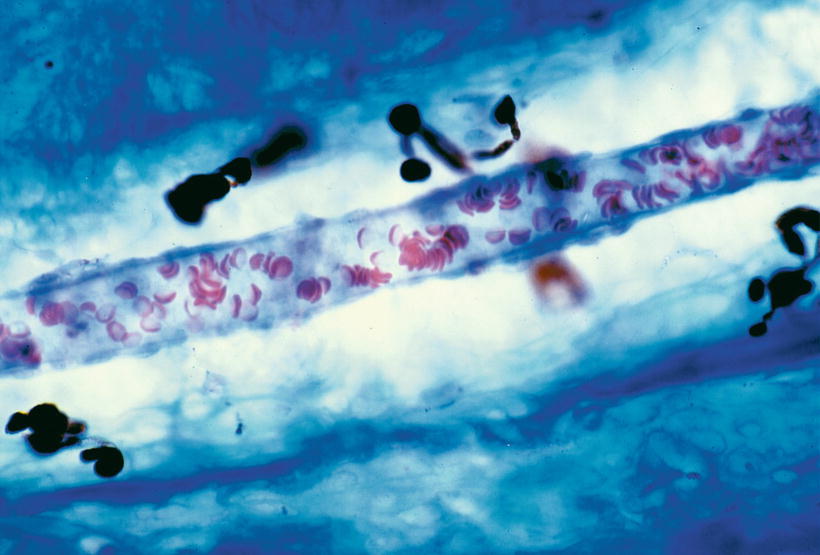
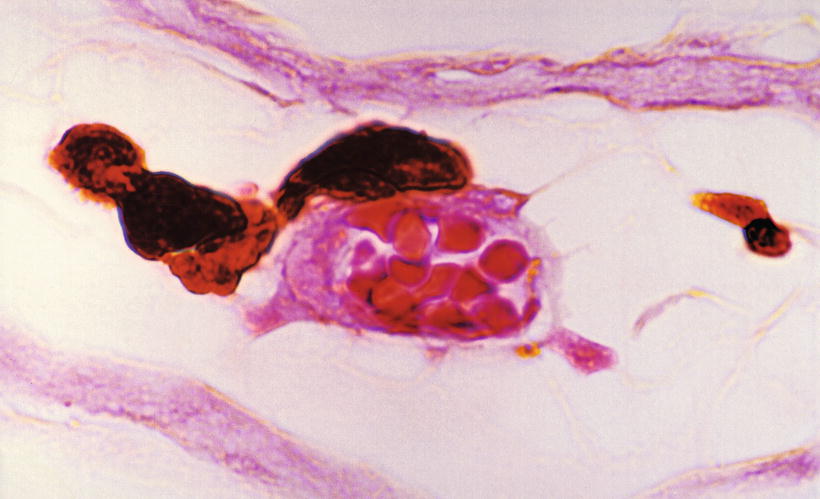
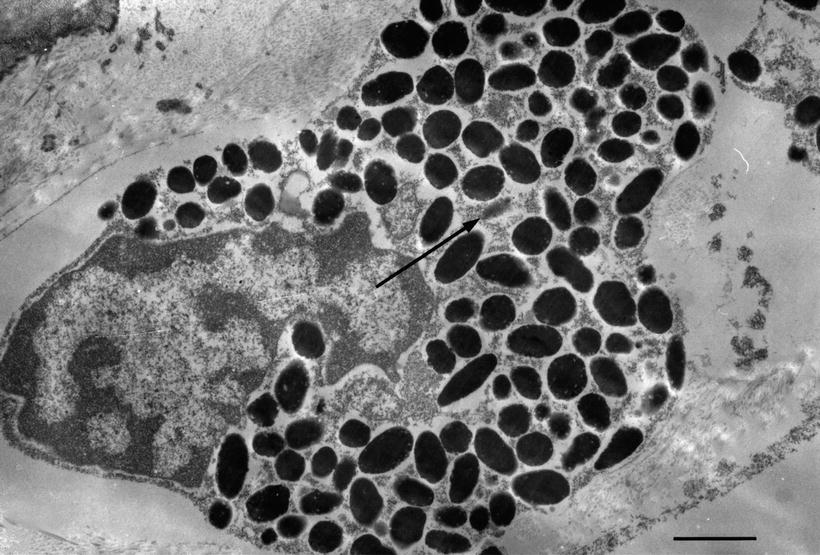
Fig. 3.6
Arterioles and venule in the septum. Arterioles (RA) run through the bony canal, while the venule (V) has no distinct canal. Azan staining (original ×6.5)
Fig. 3.7
Arteriole in the vascular canal of the septum. The diameter of the arteriole is 15 μm, while that of the vascular channel is 55 μm. Pigment cells are present in the canal. Seventy-one-year-old woman, Azan staining (original ×40) [1]
Fig. 3.8
Pigment cells adhering to the arteriole. Eighty-one-year-old woman, HE staining (original ×100) [1]
Fig. 3.9
Ultrastructure of a pigment cell. Melanosomes and premelanosomes (arrow) are observed. Sixty-one-year-old man (original ×12,000), scale: 1 μm [1]
Melanocytes are numerous in the human modiolus and their function is believed to be related to vasomotor control [2]. It has also been shown that drugs accumulate on inner ear melanin, confirmed by autoradiographic studies of lidocaine and its derivatives in rats [3].
Franz et al. [4] investigated melanocytes in the guinea pig modiolus using an electron microscope. Two different melanocyte types were observed. Type A was characterized by having spherical, uniformly pigmented stage IV melanosomes. No signs of melanogenesis were seen in this perivascular pigment cell form. Type B was very rarely observed and was characterized by fusiform to oval melanosomes in various stages of maturation. Connective tissue cells containing fusiform to oval melanosomes enclosed in membrane-bound vacuoles were frequently found. The connective tissue of the cochlear plexus seems to be derived from the leptomeninges.
To examine communication between spaces in the modiolus and the spiral ligament, Suzuki and Nomura [1] injected India ink under pressure into the modiolus. They found that India ink passed through the canal or channel of the interscalar septum to the upper spiral ligament. The bony channel ends at the margin of the interscalar septum. However, there is some continuation of the channel to the upper spiral ligament. The perivascular space around the radiating arteriole is constructed of loose connective tissue without bony tissue (Fig. 3.12). India ink was observed around the radiating arterioles (Figs. 3.10, 3.11, and 3.12), indicating that the perivascular channel is likely filled with cerebrospinal fluid (CSF).
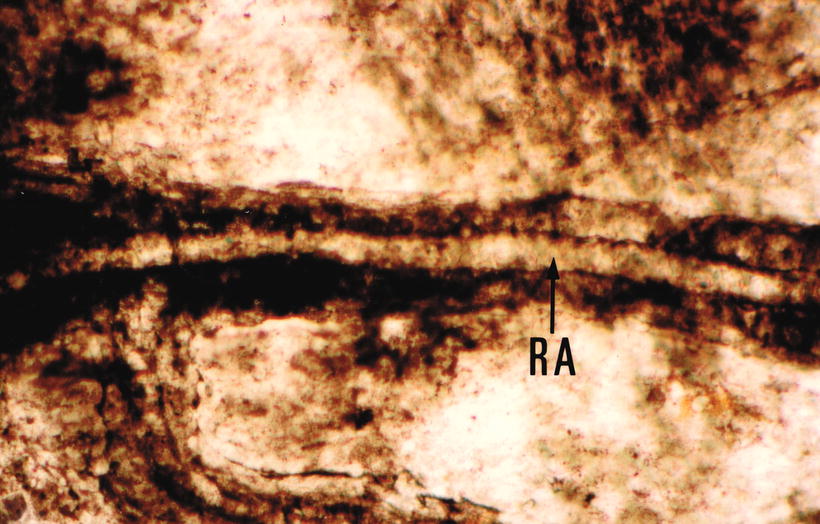
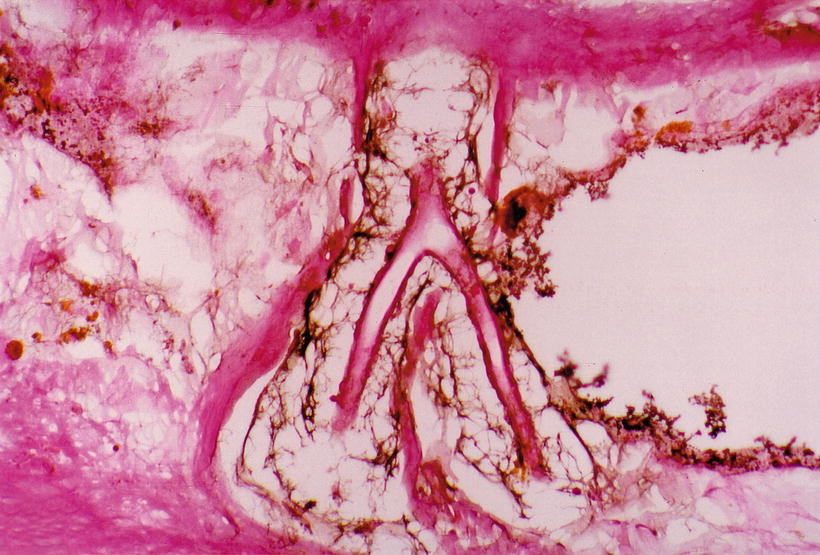
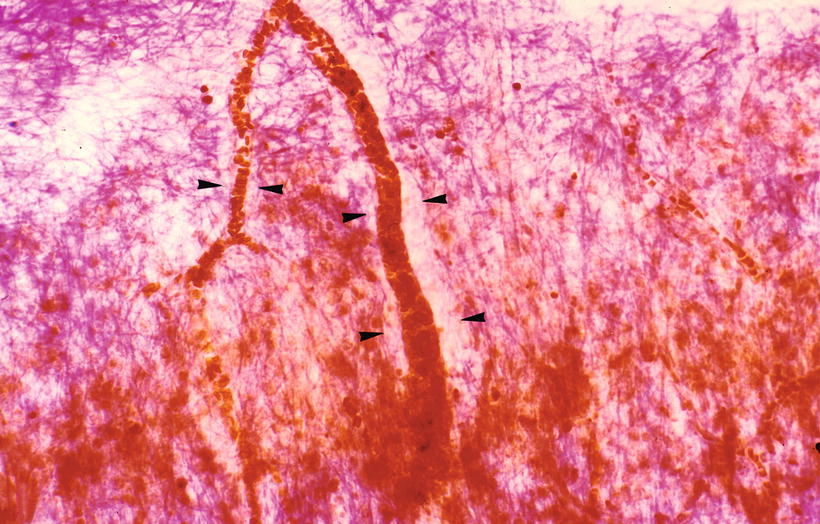
Fig. 3.10
India ink injected into the modiolus enters the vascular canal. RA radiating arteriole Sixty-eight-year-old man (original ×16) [1]
Fig. 3.11
India ink reaches the perivascular space of the upper spiral ligament. The space is surrounded by a thin bony wall. Sixty-one-year-old male, HE staining (original ×16) [1]
Fig. 3.12
Space is present around the radiating arterioles in the upper spiral ligament (arrow heads). There is no bony wall. Eight-four-year-old man, van Gieson staining (original ×16) [1]
The human modiolus facing the scala tympani has a thin and porous bony wall, and perilymph seems to pass freely through it (Fig. 3.13). The perivascular space may be involved in the formation and reabsorption of perilymph.
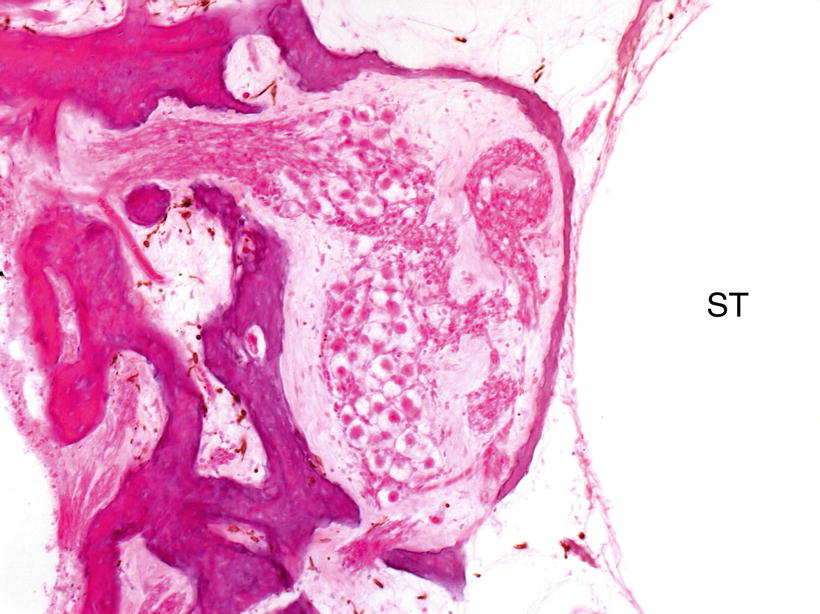
Fig. 3.13
Thin bony wall of the scala tympani of the human cochlea. Fluid communication between perilymph and cerebrospinal fluid in the modiolus is suggested. ST scala tympani
There are strand-like, atrophied vessels in the canals of the interscalar septum. Avascular canals are also observed (Figs. 3.14 and 3.15). The avascular canals may result from the disappearance of atrophied blood vessels. In the human inner ear, vascularization in the fetus is very dense, and in the infant, somewhat less so. A gradual involution of blood vessels takes place postnatally and continues with maturity and aging [5].
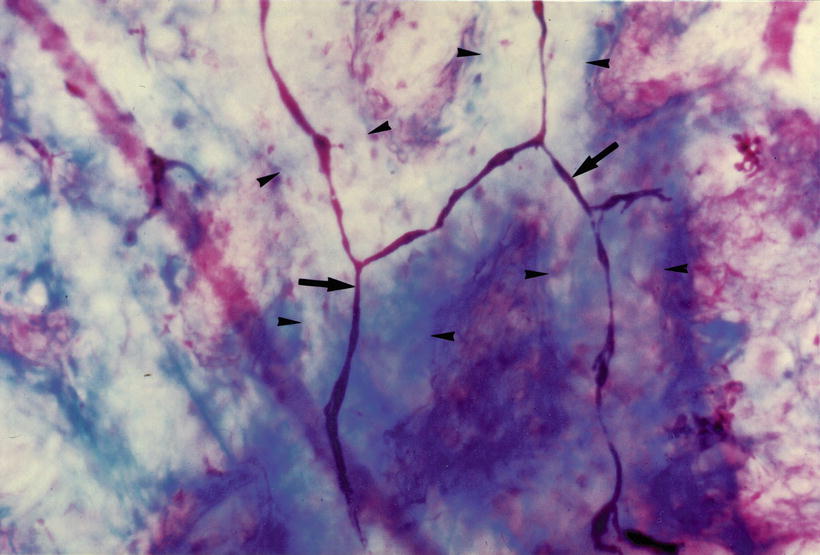
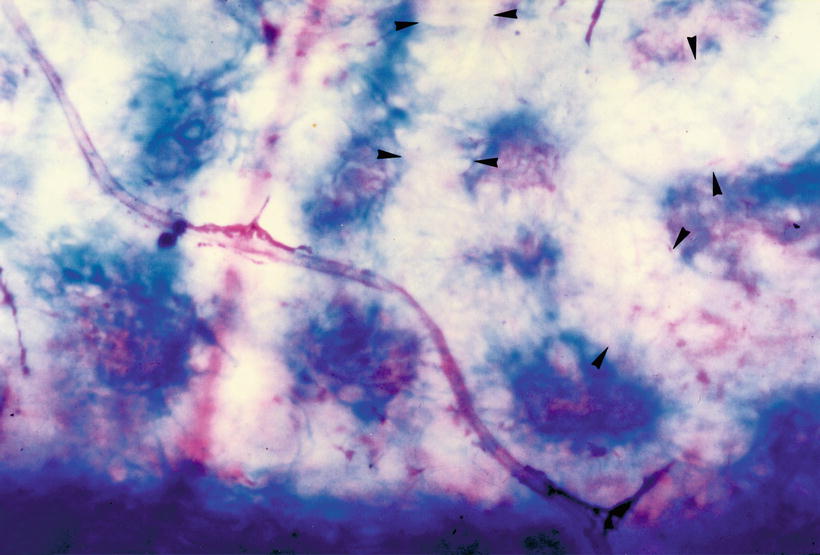
Fig. 3.14
Involution of blood vessel (arrows). Arrowheads indicate vascular channels. Eighty-three-year-old woman, Azan staining, (original ×16) [1]
Fig. 3.15
Avascular channel (arrowheads). Eighty-three-year-old woman, Azan staining (original ×16) [1]
3.1.2 Capillaries of the Membranous Cochlea
To study the blood supply to the organ of Corti, Lawrence [6] inserted a probe approximately 25 μm long into a guinea pig’s cochlea through a tiny hole in the cochlear wall to sever the arteriole going to the spiral ligament and stria vascularis. Loss of the arteriole resulted in degeneration of the spiral ligament, stria vascularis, and part of the spiral prominence, but had no effect on the organ of Corti. However, when Lawrence severed the arteriole in the modiolus going to the limbus and basilar membrane, hair cells were lost but the stria vascularis and spiral prominence remained healthy, indicating that capillary loops beneath the basilar membrane serve as the source of nutrients for the organ of Corti.
Capillary networks of the membranous labyrinth have been observed by injecting dyes, such as India ink and Prussian blue. Histochemistry is also useful for easy demonstration of the networks. Alkaline phosphatase staining is a histochemical method that can be used to demonstrate capillary networks in humans [7]. Burston’s method for alkaline phosphatase staining may be used to study the membranous labyrinth as follows:
Fresh tissue is necessary for this histochemical study. After dissection, the tissue is soaked in the staining solution overnight at 4 °C. After washing in tap water, the tissue is fixed with formalin. After decalcification and dehydration, and mounting, the stained tissue is examined and trimmed under a microscope as a surface specimen before mounting.
In this staining method, alkaline phosphatase liberates naphthol compound from its substrate, for example, naphthol AS-MX phosphate. The naphthol compound is subsequently coupled to a diazonium salt such as Fast Blue RR to produce a blue azo dye at the site of enzyme activity.
Blood vessels in the spiral ligament and stria vascularis are stained blue1 or red,2 depending on the diazonium salts used. Arterioles and venules are equally stained in humans. However, there are some species differences in staining results. Arterioles and capillaries are strongly stained in the cochlea of guinea pigs, but the venules, such as those in the lower spiral ligament, are weakly stained.
1 Blue stain | |
Naphthol AS-MX phosphate | 1.0 mg |
Dimethylformamide | 0.4 mL |
Distilled water | 12.0 mL |
0.2 M propanediol buffer at pH 8.6 | 7.6 mL |
Fast Blue RR | 20.0 mg |
2 Red stain | |
Naphthol AS-TR phosxphate | 5.0 mg |
N-dimethyl formamide | 0.5 mL |
0.05 M tris buffer as pH 8.3 | 50.0 mL |
Fast Red Violet LB | 40.0 mg |
Stained blood vessels of the stria vascularis and spiral ligament can be observed as surface specimens (Figs. 3.16 and 3.17). Strial capillaries will disappear in strial atrophy.
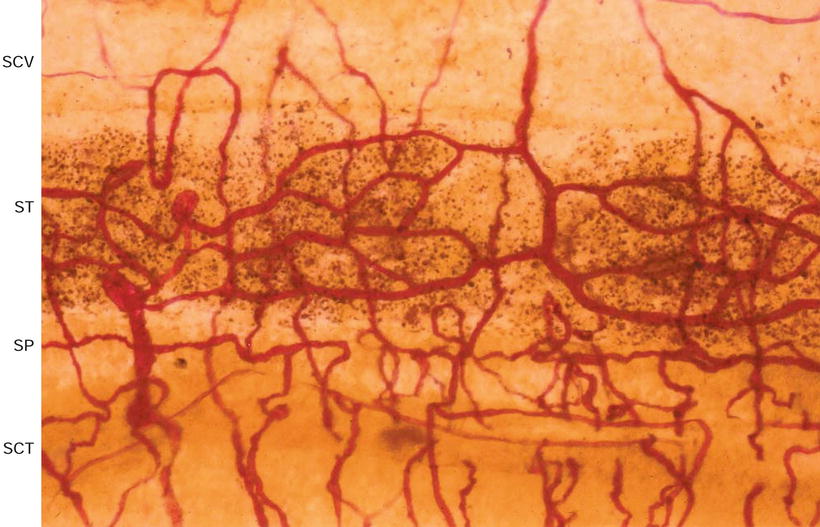
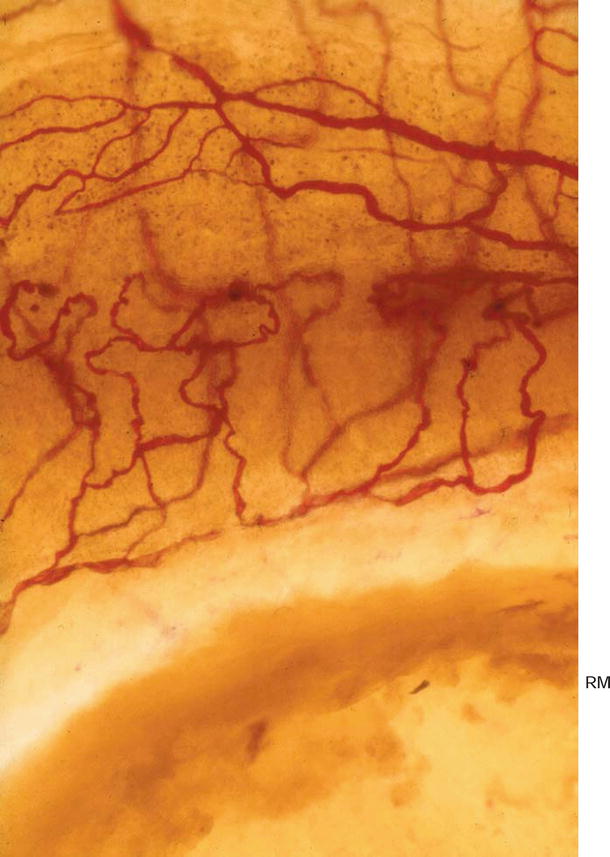
Fig. 3.16
Blood vessels of the stria vascularis and spiral ligament. SCV scala vestibuli, ST stria vascularis, SP spiral prominence, SCT scala tympani. Eighty-six-year-old woman, alkaline phosphatase staining (Burston), (original ×6.5)
Fig. 3.17
Blood vessels of the stria vascularis and spiral ligament adjacent to the round window membrane (RM). Eighty-six-year-old man, alkaline phosphatase staining (Burston), (original ×6.5)
Branches of the radiating arterioles run in the spiral ligament to join the venules as arteriovenous arcades or join the vessel of the spiral prominence (Fig. 3.18). The spiral prominence and root cells are endowed with rich enzymatic activities (Fig. 3.19) [8].
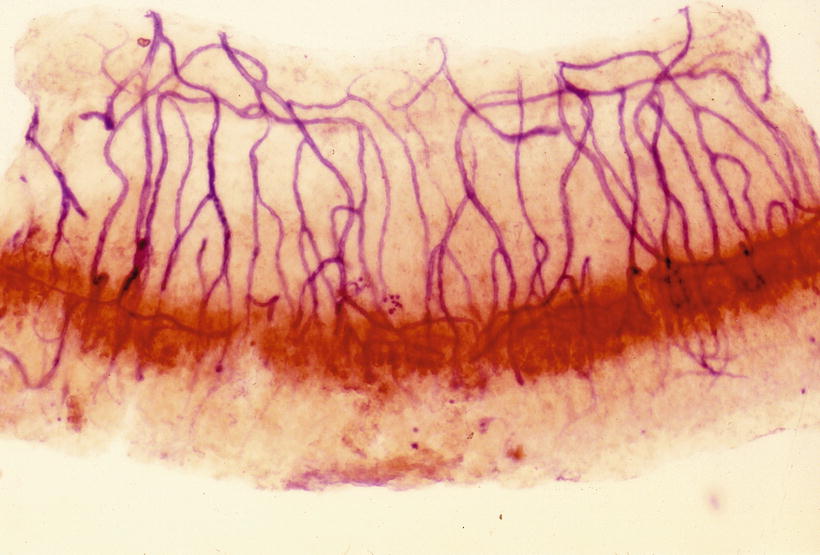
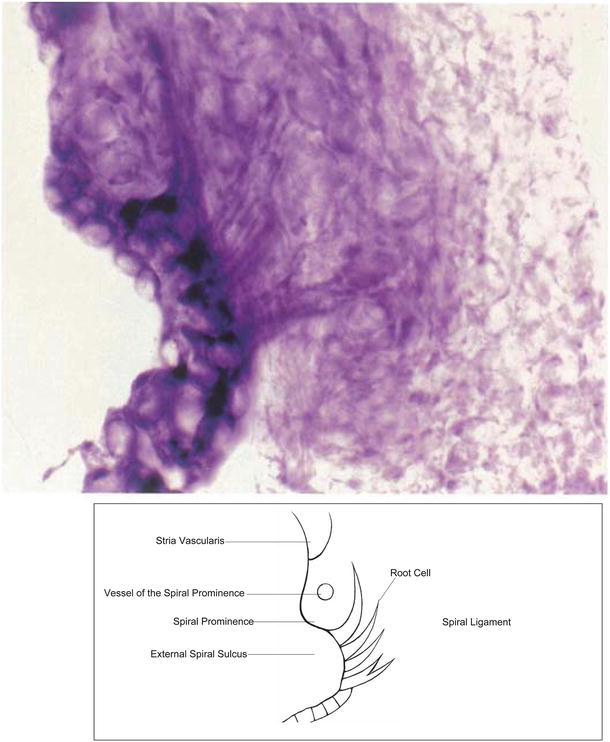
Fig. 3.18
Arteriovenous arcades in the spiral ligament and the root cells. Butylcholinesterase activity is present in the root cell. The blood vessels were stained to demonstrate alkaline phosphatase. The venules in the lower spiral ligament shows weak activity of alkaline phosphatase. The stria vascularis was removed. Guinea pig, double staining
Fig. 3.19
NADH diaphorase activity in the root cells. Guinea pig. NAD Nicotinamide adenine dinucleotide
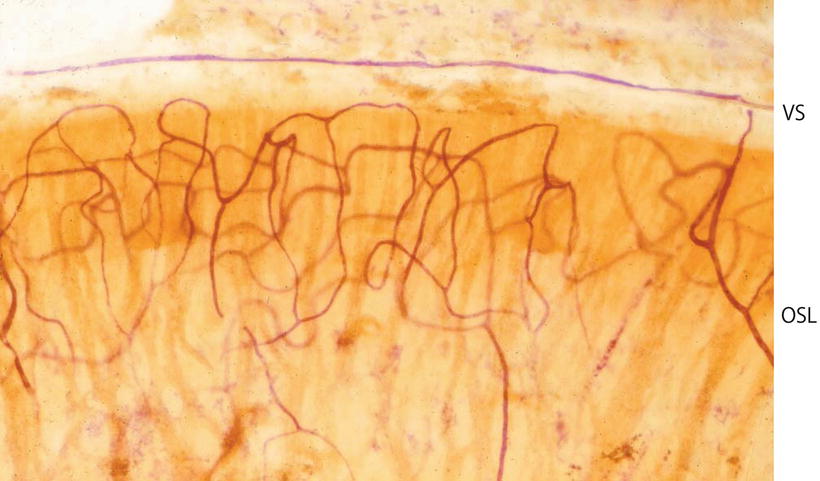
Blood vessels in the osseous spiral lamina, organ of Corti, and limbus spiralis can also be clearly visualized [9, 10] (Figs. 3.20, 3.21, and 3.22). Capillaries in these structures are principally loops arranged in a coronal pattern. The loops run from the modiolus laterally in the osseous spiral lamina and underneath the basilar membrane, then return to the modiolus. It is interesting to note that no outer spiral vessel (vas spirale, the spiral vessel under the tunnel of Corti) is present beneath the organ of Corti in cats (Fig. 3.23).
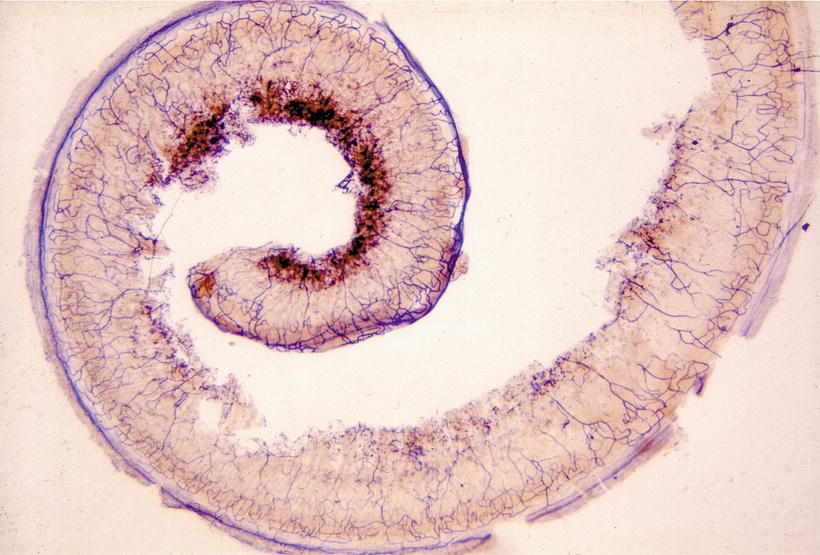
Fig. 3.20
Capillaries in the osseous spiral lamina with the organ of Corti of the whole cochlea. The capillary networks are dense in the osseous spiral lamina of the upper turn in comparison with the lower basal turn. Seventy-nine-year-old man, alkaline phosphatase staining (Burston), (original ×2.5) [9]