Aqueous humor
Trabecular meshwork
Optic nerve
Systemic (serum/plasma)
ECM elements
CD44
↓ [13]
Cochlin
↑ [14]
Chondroitin sulfate
↑ [15]
Collagen type IV
nc [16]
Elastin
↑ [17]
↑ [18]
Fibronectin
nc [19]
nc [16]
Hyaluronic acid
↓↓ [20]
↓↓↓↓ [15]
↓ [21]
GAGase-resistant material
↑↑↑↑ [15]
Tenascin
↑ [22]
Thrombospondin-1
↑ [23]
ECM remodeling enzymes and inhibitors
MMP-1
↑ [24]
↑ [25]
MMP-3
nc [26]
↑ [24]
↑ [25]
MT1-MMP
↑ [27]
TIMP-1
nc [26]
↑ [24]
TIMP-2
↑ [28]
Table 27.2
Cell signaling changes in POAG
Aqueous humor | Trabecular meshwork | Optic nerve | Systemic (serum/plasma) | |
|---|---|---|---|---|
Angiopoietin-like 7 | ↑↑↑ [29] | |||
Aquaporin-9 | ↓ [30] | |||
Erythropoietin | ↑ [31] | |||
Endothelin-1 | ↑ [32] | |||
Hepatocyte growth factor | ↑ [34] | |||
Phospholipase A2 | ↑ [35] | |||
Plasminogen activator inhibitor-1 | ↑↑↑ [36] | |||
Soluble CD44 | ||||
Thymulin | ↑↑↑ [39] | |||
Vascular endothelial growth factor | ↑ [40] | ↑ [41] |
Table 27.3
Oxidative stress and senescence changes in POAG
Aqueous humor | Trabecular meshwork | Optic nerve/retina | Systemic (serum/plasma/red blood cells) | |
|---|---|---|---|---|
Acetylcholinesterase | ↑ [42] | |||
Asymmetric dimethylarginine | ↑ [43] | |||
3-α-Hydroxysteroid dehydrogenase | ||||
Ascorbic acid | ↑↑↑ [46] | |||
Citrate | ↓ [47] | |||
Cortisol | ↑ [48] | |||
Diadenosine tetraphosphate | ↑ [44] | |||
Dimethylarginine | ↑ [43] | |||
ELAM-1 | ↑ [49] | |||
Fatty acid | ||||
Eicosapentaenoic | ↓ [50] | |||
Choline plasmalogens | ↓ [51] | |||
Docosahexaenoic | ↓ [51] | |||
Ω-3 | ↓ [51] | |||
Glutathione | ↑↑↑ [52] | ↓ [53] | ||
Nitric oxide | ↑ [54] | |||
Nitric oxide synthase | ↑ [55] | |||
NF-κB | ↑ [56] | |||
HIF-1α | ↑ [57] | |||
Senescence-associated β-galactosidase | ↑ [58] |
Table 27.4
Innate and adaptive immunity changes in POAG
Aqueous humor | Trabecular meshwork | Optic nerve/retina | Systemic (serum/plasma) | |
|---|---|---|---|---|
Innate immunity | ||||
Receptors | ||||
TLR 2 | ↑↑ [59] | |||
TLR 4 | ↑↑↑ [59] | |||
TLR 7 | ↑ [59] | |||
TLR 8 | ↑ [59] | |||
Associated proteins | ||||
MyD88 | ↑ [59] | |||
CD44 | ↑ [13] | |||
Ligands (pathogen-associated molecular patterns) | ||||
Hsp 60 (bacterial) | ||||
H. pylori | ↑↑↑ [62] | |||
Heat shock proteins | ||||
Hsp 27 | ↑↑↑↑ [64] | ↑ [65] | ||
αB-crystallin | ↑ [65] | |||
Hsp 60 | ↑↑↑↑ [59] | ↑ [65] | ||
Hsp 70 | ↑↑↑↑ [59] | ↑ [65] | ||
Serum amyloid A | ↑↑ [66] | |||
Adaptive immunity | ||||
Cytokines | ||||
IL-2 | ↑ [67] | |||
IL-4 | ↑↑ [68] | |||
IL-8 | ||||
IL-12 | ↑↑ [68] | |||
TGF-β2 | ||||
TNF α | nc [73] | ↑ [25] | ↓ [68] | |
Autoantibodies | ||||
Anti-phosphatidylserine | ↑ [74] | |||
Fodrin | ↑ [75] | |||
Glial fibrillary acidic protein | ↓ [74] | |||
Glutathione S-transferase | ↑ [76] | |||
Myelin basic protein | ↑ [74] | |||
Neuron-specific enolase | ↑ [77] | |||
Retinaldehyde-binding protein | ↑ [74] | |||
Retinal S-antigen | ↑ [78] | |||
Vimentin | ↑ [65] | |||
γ-Enolase | ↑ [74] | |||
14-3-3 | ↓ [74] | |||
Monoclonal gammopathy | ↑ [76] | |||
ECM Changes in POAG
The ECM and TM endothelium are essential for maintenance of the normal aqueous humor outflow [79]. In the TM of POAG eyes, excessive, abnormal accumulations as well as decreases of other ECM materials (Table 27.1) have been documented [80, 81]. The ECM produced by the cells is composed of multidomain macromolecules such as collagens, cell-binding glycoproteins, and proteoglycans that link together to form a structurally stable composite. It is now well documented that ECM is a dynamic entity determining and controlling the behavior and biologic characteristics of the cells [82].
One key component of the ECM in the TM is proteoglycans which are macromolecules consisting of a core protein to which glycosaminoglycan side chains are covalently attached. This class of molecules has been implicated in the maintenance of resistance to aqueous humor outflow. In the TM tissue, proteoglycans form gel-like networks that may function as a gel filtration system. The major types identified include chondroitin, dermatan, and heparan sulfate proteoglycans that may represent decorin, biglycan, versican, perlecan, and syndecan [82, 83].
The relative amounts of each type of glycosaminoglycan in the TM tissue have been determined [15, 80]. Hyaluronic acid and chondroitin–dermatan sulfates are the major constituents, with heparan sulfate and keratan sulfate present in smaller amounts. A depletion in hyaluronic acid and an accumulation of chondroitin sulfates and undigestible glycosaminoglycan material have been associated with POAG conditions [15]. Both chondroitin sulfate and hyaluronic acid have been shown to contribute to flow resistance and influence flow rate in vitro [80]. The flow rate was decreased when hyaluronic acid and chondroitin sulfate were used at POAG concentrations. If glycosaminoglycan chain biosynthesis is distributed by sodium chlorate or β-xyloside, outflow facility increases in perfusion cultures [81]. Of note, the level of an ectodomain fragment of hyaluronic acid receptor CD44 (sCD44) was found to be elevated (Table 27.2) in the aqueous humor of POAG patients [37], and the concentration was highly correlated with the clinical visual field loss which characterizes POAG. sCD44 is internalized in TM cells and localizes in part to mitochondria [84]. sCD44 is cytotoxic to TM cells [85].
Fibronectin, laminin, vitronectin, and matricellular proteins that include tenascin and thrombospondin-1 have been localized in the TM [82]. These glycoproteins are crucial in biologic processes such as cell attachment, spreading, and cell differentiation [82]. Overexpression of fibronectin, laminin, and collagen type IV results in a decrease in the TM cell monolayer permeability [86, 87]. In addition, the expression of thrombospondin-1 has been shown to be increased [23] in the TM of POAG eyes (Table 27.1).
Elastin is localized to the central core of sheath-derived plaques or elastic-like fibers in the TM [86]. Fibrillin-1, a component of microfibrils, is found in both the core and the surrounding sheath of the elastic-like fibers. Fibrillin-1 and collagen type VI are also constituents of long-spacing collagens in the TM [86, 87]. In trabecular lamellae and in juxtacanalicular tissue, accumulation of long-spacing collagens and sheath-derived plaques has been documented in POAG and aged eyes [86, 88]. The cochlin deposits in the glaucomatous TM (Table 27.1) appear to increase with age and are associated with proteoglycans. Such deposits have been proposed to contribute to the increase of ECM resistance to outflow and the POAG pathology [12].
The ECM is constantly modified by the surrounding cells through enzymes such as matrix metalloproteinase (MMP) family member and inhibitors such as tissue inhibitors of matrix metalloproteinase (TIMPs) found in the TM [83]. Ongoing ECM turnover, initiated by MMPs, appears to be essential for maintenance of the aqueous outflow homeostasis. MMP-3, and possibly also MMP-9, may be responsible for the efficacy of laser trabeculoplasty, a clinically useful alternative treatment to reduce IOP in patients with glaucoma [83, 86]. Addition or induction of MMP-3 in perfused human anterior segment organ cultures increases, whereas blocking the endogenous activity of the MMPs in the TM reduces, the aqueous humor outflow facility [83].
The ECM in the TM may also be remodeled in response to exogenous stimuli such as glucocorticoids and oxidative stress [82, 86]. Mechanical stretch caused an increase in MMP-1 and MMP-3 activities and alteration of ECM molecules including proteoglycans and matricellular proteins [89]. The ECM is modulated by cytokines. The most studied cytokine in the TM is transforming growth factor-beta (TGF-β). A higher than normal level [73] of TGF-β2 was found in the aqueous humor of patients with POAG (Table 27.4). In TGF-β2-perfused organ cultures, focal accumulation of fine fibrillar extracellular material was observed in TM tissues. Furthermore, TGF-β2 perfusion reduced outflow facility and elevated IOP [90], and adenoviral gene transfer of active TGF-β2 elevated IOP and reduced outflow facility in rodent eyes [91]. These results suggest that the increased TGF-β2 level in the aqueous humor may be related to the pathogenesis of glaucoma. Other cytokines such as tumor necrosis factor-α (TNF-α) that is increased in the optic nerve head of POAG (Table 27.4) also modulate the ECM, probably via regulation of MMP and TIMP expressions [82, 92].
Signaling Molecules in POAG
The TM and optic nerve utilize local and probably systemic cell signaling pathways to maintain cell viability. Locally in the TM, the Rho family of small guanosine triphosphatases (GTPase) has been shown to be of vital importance in the outflow system [93–95]. Sphingosine-1-phosphate and endothelin-1 activate Rho/Rho kinase signaling pathway decrease the aqueous humor outflow facility [93–95]. Endothelin-1 has been reported to be increased in the aqueous humor [32] and blood [33] of POAG patients (Table 27.2).
The aqueous humor also modulates TM cell signaling. It contains albumin as a major constituent. Other components encompass hydrogen peroxide (H2O2), ascorbic acid, cytokines such as TGF-β, hepatocyte growth factor and vascular endothelial growth factor (VEGF), and molecules including MMPs, proteinase inhibitors, sCD44, and hyaluronic acid [82]. In both POAG aqueous humor and Alzheimer’s cerebrospinal fluid [96], hepatocyte growth factor [34] and VEGF [40] increase, indicating modulation of these fluids in the disease process. Increased levels of angiopoietin-like 7, aquaporin-9, erythropoietin, plasminogen activator inhibitor-1, sCD44, interleukin-2, phospholipase A2, thymulin (Table 27.2), glutathione, ascorbic acid (Table 27.3), and TGF-β2 (Table 27.4) but, significantly, a decreased level of hyaluronic acid (Table 27.1) have been reported in the aqueous humor of POAG eyes. Of note, the decrease in aqueous hyaluronic acid concentration parallels that in the TM, and the increase in sCD44 (the ectodomain portion of the CD44 receptor) also parallels the CD44 receptor increase in the TM (Table 27.1). Only sCD44 has been tested in rodents; adenoviral gene transfer of sCD44 resulted in a sustained increase in IOP in mice [97]. The sCD44 found in the POAG aqueous humor is hypophosphorylated [98]. The hypophosphorylated form has high cytotoxicity and low hyaluronic acid-binding affinity and is suggested to represent a pathophysiologic feature of the disease process [98].
Stress and Aging in POAG
TM cellularity is reduced with aging [99, 100]. Morphologic studies have also revealed thickened basement membranes and accumulation of sheath-derived plaques and long-spacing collagens in the TM of aged eyes [90]. The number of senescent cells which stain positive for senescence-associated β-galactosidase is increased (Table 27.3) in the TM of POAG eyes [56], supporting further that POAG is an age-related disease [82, 101]. Oxidative damage and stress (Table 27.3) have also been implicated to contribute to the morphologic and physiologic alterations in the aqueous outflow pathway in aging and glaucoma [102]. The TM is subjected to chronic oxidative stress [86]. Superoxide dismutase, an enzyme involved in the protection against oxidative damage, has been shown to decline with age in human TM tissues [101]. TM cells also synthesized a specific set of proteins, such as αB-crystallin, that may act as molecular chaperones to prevent oxidative or heat shock protein damage [41]. Markers of oxidative damage [102, 103], abnormalities in mitochondrial DNA [104], and diminished blood levels of oxidant scavenger glutathione [52] are found in POAG patients. It appears that oxidative stress that exceeds the capacity of TM cells for detoxification is involved in damaging the cells and alteration of the aqueous humor outflow, leading to increased IOP. Moreover, stress-/aging-related changes also occur in the red blood cell phospholipid composition which may alter red blood cell flexibility in POAG [51], causing reduced ocular blood flow and optic nerve axon loss. Linear regression analysis suggests that a decrease in phosphatidylcholine-carrying docosahexaenoic acid (DHA, an omega-3 fatty acid) occurs years before the onset of POAG. This underscores the notion that aging, and specifically membrane lipid composition, is a risk factor in POAG [51]. In a longitudinal study, a decrease in phospholipid composition correlates with the development of Alzheimer’s disease [105]. There is, however, a possible correction for the change in lipid composition of membranes. One effect of dietary omega-3 is downregulation of Toll-4 innate immune receptors [106], whereas supplement-free fatty acids upregulate Toll-4 [107].
Innate and Adaptive Immunity in POAG
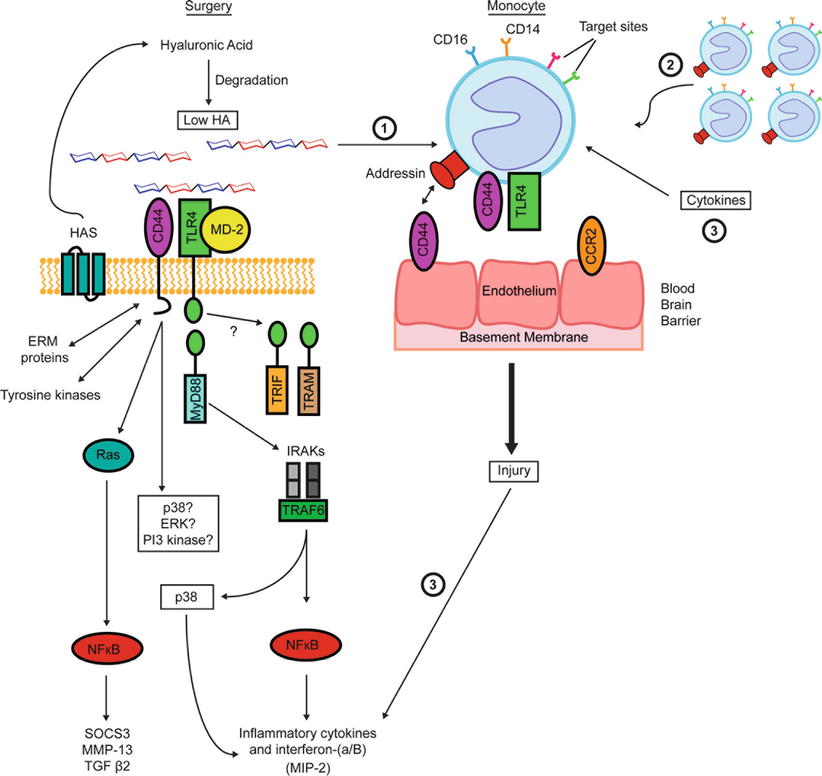
The immune system is actively involved in the POAG disease process. Innate immunity is the first line of defense and is rapidly activated by surgery (see Fig. 27.1), leading to NF-kB activation responsive genes [108]. Toll-4 receptor recognizes low molecular weight hyaluronic acid fragments and many others [108]. Significantly, a number of Toll-like receptors are increased in the retina of POAG patients (Table 27.4). Upregulation of acute stress response protein amyloid in TM of POAG patients [66] further supports the notion that POAG has ocular altered protein expression. Treatment of cultured human TM cells with recombinant serum amyloid A affects gene expression, including a 22-fold upregulation of interleukin-8 [66]. Toll-like receptors are expressed on monocytes, dendritic cells, neutrophils, mucosal epithelial cells, and endothelial cells [109]. Notably, low molecular weight hyaluronic acid activates monocyte expression. Monocytes from the blood extravasate through the blood vessel walls using the multifaceted CD44, the homing receptor for leucocytes. Once monocytes arrive in the wound site, monocytes differentiate into macrophages. Upon activation, the macrophages induce new ECM and stimulate the local cells to re-epithelize the wound. Figure 27.1 depicts three pathways in which monocytes are activated: (1) low molecular weight hyaluronic acid, (2) cytokines and chemokines, and (3) recruitment of other monocytes. The monocyte activation creates positive feedback loop leading to extensive scar tissue and aberrant wound healing. See Fig. 27.1. One idea that we are exploring is that the expression of CD44 message and its numerous variants are changed in POAG. Although CD44 expression on keloid cells is similar to normal dermal cells [110], keloid tissue exhibits a different miRNA profile [111, 112] and increased hyaluronic acid [110]. The 3′ untranslated region of CD44 is very active in miRNA binding and influences disease progression [113]. The overexpression of sCD44 in the aqueous of African-Americans with POAG [37] is important since CD44 contributes to inflammation and fibrosis in response to injury. CD44 promotes the activation of TGF-β via an MMP-dependent mechanism leading to fibroblast migration and recruitment [114]. We have used flow cytometry to characterize monocyte activation. See Fig. 27.2. Recently, three monocyte subset populations were identified as classical (CD14++CD16−), intermediate (CD14++CD16+), and nonclassical (CD14+ CD16++) [115]. In healthy individuals the classical monocytes represent ~90 % of monocytes, while the intermediate and nonclassical comprise the other 10 [116]. The relative concentrations of intermediate and nonclassical monocytes have been implicated in a number of diseases, such as HIV-1 infection [117], cancer [118], and stroke [119]. Notably, monocyte profile predicts the clinical course and prognosis human stroke [119]. Monocyte subtypes have already been identified in Eales disease [120], but to our knowledge have not been examined in POAG. If monocyte subtypes are different in POAG from normal patients, monocyte subtypes may provide a reliable biomarker of the disease as well as information about patient’s status after glaucoma-associated surgery. Interestingly, a recent paper identified that radiation treatment has been found to prevent glaucoma in a mouse model by disrupting monocyte migration [121]. One particular element that was disrupted was l-selectin, which, when activated, aids in trans-endothelial cell migration. The intermediate monocytes (CD14++CD16+) overexpress l-selectin, which might implicate this monocyte subtype as a prime facilitator of glaucoma [122]. If these results are supported, patients undergoing surgery might benefit from having their monocyte subtypes examined using flow cytometry.