Binocular Vision Adaptations in Strabismus
Marshall M. Parks
Strabismus patients with established binocular vision before the onset of misaligned eyes may adapt their binocular vision to conform to the deviation of their eyes. The adaptations are in response to the annoying symptoms caused by the persistent functioning of normal binocular vision after acquiring the strabismus. Before discussing the adaptation, a brief review of normal binocular vision is in order.
BINOCULAR VISION
Binocular vision is the cortical integration of similar images on each retina into a unified perception. Binocular vision is composed of three independent components and is contained in two separate reflexes. The components of binocular vision are simultaneous perception, fusion, and stereopsis. The distinct reflexes of macular and extramacular binocular vision result in the duality of binocular vision. The many differences between the macular and extramacular binocular vision reflexes are discussed in Chapter 5.
Pertinent to this chapter on the adaptations of binocular vision in strabismus and the most important difference between macular and extramacular fusion reflexes is the absence of any adaptation by the macular fusion reflex in the strabismic patient. The reflex is impossible to adapt to the newly acquired strabismus because it ceases to function the moment the eyes deviate from being straight. For an adaptation of the binocular vision reflex to occur, the reflex must continue functioning during strabismus, which causes annoying symptoms that are eliminated by the adaptation. Only the extramacular fusion reflex continues functioning after the onset of the strabismus. Hence, the adaptations of binocular vision in strabismus are restricted to the extramacular fusion reflex.
Binocular vision is an acquired reflex that normally develops during the first 3 to 4 months of life. Its development demands certain requisites. The infant must be capable of seeing with each eye, and both eyes must be aligned with one another, permitting similar retinal images to project onto corresponding retinal areas during the critical period for binocular vision development, which extends to approximately 2 years of age. Patients with congenital or very early onset strabismus do not receive the essential stimulation from similar images projecting onto corresponding retinal areas; consequently, their binocular vision reflexes do not develop. Absence of binocular vision is elicited by sensory tests yielding responses that prove the absence of simultaneous perception of images on each retina (Chapter 9).
Patients1,2 without binocular vision experience none of the untoward visual symptoms that are associated with newly acquired strabismus in those who have developed binocular vision. Their only visual handicap is absence of stereopsis, which they are as unable to comprehend as the colorblind person is unable to comprehend a deficiency in color perception.
Despite the absence of binocular vision, if the attention directed to the object of regard results in fixating first with one eye for a moment and then fixating with the other eye for the next moment, such alternate fixation ensures normal monocular macular function in each eye. However, if the attention to the object of regard was fixated exclusively with one eye during the amblyogenic age, amblyopia will be apparent in the unused eye.
VISUAL SYMPTOMS IN STRABISMUS
Once the extramacular fusion reflex has developed, annoying visual symptoms are apparent when the eyes deviate from being straight or when placing a 15Δ prism before one eye. Viewing with the strabismic eyes, or through the prism before straight eyes, precludes the opportunity for similar retinal images to project onto corresponding retinal areas. The similar images are now seen doubled in space (diplopia), along with an overlay of dissimilar images appearing to be superimposed in the same place in space (visual confusion).
DIPLOPIA
Diplopia results from the simultaneous perception of the images of the object of regard projecting onto noncorresponding retinal areas of the strabismic eyes. The simultaneous perception of the two similar images under these circumstances yields the impression that the object of regard is simultaneously located at two points in space (Fig. 1).
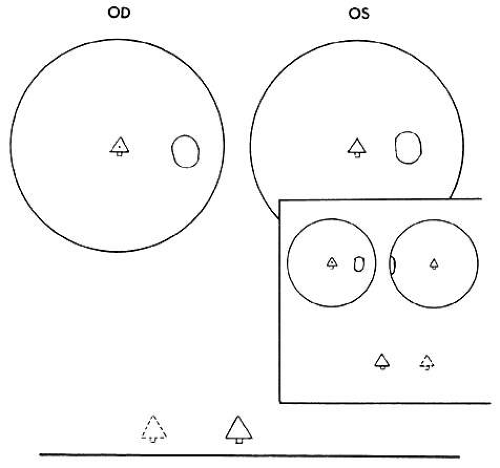 Fig. 1. Diplopia results from simultaneous perception of similar images of the object of regard, that is, a Christmas tree, projecting onto noncorresponding retinal areas. Esotropia causes homonymous diplopia; clear image on macula of right eye is illustrated by solid lines, blurred image on nasal retina of left eye is illustrated by broken lines. The illustrated diplopic images are displayed according to the patient’s drawing of them. Inset. Heteronymous diplopia in exotropia. (From Symposium on Strabismus, Transactions of the New Orleans Academy of Ophthalmology. St Louis: CV Mosby, 1971.) |
The size of the retinal area in one eye that corresponds to a retinal point in the other eye rapidly expands for retinal loci increasingly peripheral to the fovea. This determines the degree of retinal image disparity that either the macular or extramacular fusion reflex find tolerable. It accounts for Panum’s visual space. Thereby, the depth of Panum’s visual space is very limited for the macular fusion reflex to function within compared with the ever-expanding depth of fusional space within which the extramacular fusion reflex operates. This explains how an esotropic deviation may be sufficiently small to allow a spatial target peripheral to the object of regard (yet within Panum’s fusional space) to project the peripheral target’s images onto corresponding retinal areas while the object of regard projects its images onto noncorresponding macular areas (Fig. 2).
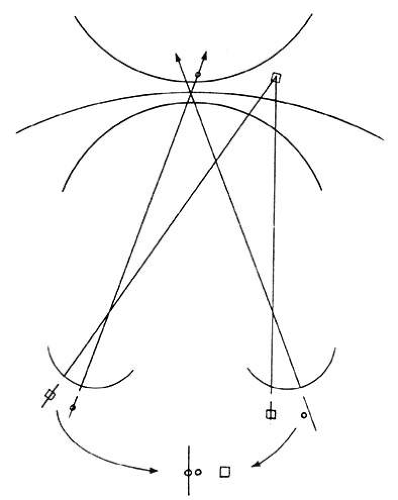 Fig. 2. The heterotropic deviation may be sufficiently small to cause the object of regard to be diplopic, but the peripheral objects displaced equidistant to the object of regard from the horopter may still be within Panum’s area. Thus it is possible for a patient with a small heterotropic deviation to have extramacular fusion in the absence of macular fusion. (From Symposium on Strabismus, Transactions of the New Orleans Academy of Ophthalmology. St Louis: CV Mosby, 1971.) |
According to Ogle,3 foveal horizontal image disparity greater than 20 minutes of arc (2/3 of a prism diopter) is not fusible. From my clinical investigation it appears that extramacular image disparity is fusible up to approximately 480 minutes of arc4,5 (8 prism diopters). The vast differences in tolerance of horizontal image disparity between macular and extramacular fusion suggest they are under the control of separate binocular vision reflexes. This reasoning is strengthened by the frequency of clinical cases that are devoid of a macular fusion reflex but have a normal extramacular fusion reflex.
The purpose of the macular fusion reflex is to produce and maintain bifixation. If the reflex is absent or intermittently shut down for any reason, the patient monofixates. Monofixation is manifest by a three-degree facultative macular scotoma present only in the nonfixating eye during binocular viewing while the intact normal extramacular fusion reflex remains within 0 to 8 prism diopters of horizontal eye alignment.
Such a large range of tolerance for horizontally disparate retinal images to be fused in the monofixator is easily demonstrated using stereopsis tests. Although the stereoacuity is poor in the monofixator compared with the bifixator,6 it does exist as long as the horizontal deviation of the eyes does not exceed 8 prism diopters. Along with the intact stereopsis, the fusional vergence amplitudes also remain normal in the monofixator as long as the horizontal deviation of the eyes does not exceed 8 prism diopters. In my experience, normal fusional vergence amplitudes and stereopsis perception occur only in normal retinal correspondence (NRC) but not in the adapted abnormal retinal correspondence (ARC).
Stereopsis can occur only if the horizontally disparate images in the monofixator’s peripheral vision are from targets within Panum’s fusional space. Although the center of the Panum’s area is scotomatized, the peripheral Panum’s area exists in small angles of deviation. To maintain this small horizontal deviation of 0 to 8 diopters (D) requires a good functioning fusional vergence reflex that can be generated by NRC but not by ARC.
VISUAL CONFUSION
Visual confusion occurs in strabismus as a result of the projection onto corresponding retinal areas of simultaneously perceived dissimilar images of two different spatial objects that normally project onto noncorresponding retinal areas. Consequently, these different objects in space that create dissimilar images are perceived as located at the same place in space. Visual confusion does not exist for all portions of the visual field. The object of regard visually perceived by the fixating eye cannot simultaneously perceive a superimposed different image projecting on the fovea of the deviating eye. From the physiologic fact of binocular vision, as stated in Chapter 5, Fig. 13, only similar images projecting on the foveas are perceived simultaneously. No binocular function results from dissimilar images projecting onto these retinal areas. Consequently, from the moment the strabismic eyes deviate the patient is spared the visual confusion of a different spatial object’s image conflicting with the object of regard. The three-degree macular scotoma in the esotropic left eye (Fig. 3) can be plotted using a binocular perimetric technique. This is a physiologic scotoma, present in all strabismic patients immediately at the moment the alignment of the eyes exceeds the threshold of 2/3 of a prism diopter. Therefore sustaining the bifixation reflex requires a very refined ocular alignment control along with an elegant level of similarity of the foveal images. The combination of these two inputs provide the essential stimuli for bifixation. The moment one or both are lacking, the bifixation reflex shuts down and a macular scotoma appears in the nonfixating eye. It is not a pathologic scotoma that evolves gradually to resolve the annoying visual symptom caused by strabismus. It is not an adaptation of the macular fusion reflex that gradually developed to eliminate visual confusion. In short, it is not a suppression scotoma.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


