, Yasuya Nomura2 and Yasuya Nomura3
(1)
The Society for Promotion of International Oto-Rhino-Laryngology, Tokyo, Japan
(2)
The University of Tokyo, Tokyo, Japan
(3)
Showa University, Tokyo, Japan
Abstract
Some viral infections are known to cause sensorineural hearing loss, which may occur suddenly as an acute single insult, or insidiously with a slowly progressive course. When hearing loss occurs during the course of systemic viral infection, it will be apparent that the infection is responsible for the hearing loss. However, hearing loss is not uncommonly categorized as being of “unknown etiology” and left undiagnosed.
To promote better understanding of viral labyrinthitis, we will present human temporal bone histopathology and animal experiment results concerning three viruses: herpes simplex virus, cytomegalovirus, and mumps virus.
In human temporal bone specimens from patients with viral labyrinthitis, the degenerated tectorial membrane often becomes rolled-up and encapsulated on the limbus spiralis or in the inner sulcus. Such findings are seen in patients with mumps and measles. In experimental herpes simplex virus labyrinthitis in guinea pigs, a rolled-up tectorial membrane with viral antigen deposition has been observed, along with bulges of varying sizes in the tectorial membrane. These bulges were found on the surface of the membrane as well as within it. With electron microscopy, virions were observed in and around the bulges. Similar findings were observed in a patient with bilateral sudden deafness. Reactivation of latent HSV infection was suspected in that case. However, not all cases of viral labyrinthitis show changes in the tectorial membrane.
Keywords
CytomegalovirusExperimental viral labyrinthitisHerpes simplex virusHuman temporal bone histopathologyMumps6.1 Herpes Simplex Virus (HSV)
6.1.1 Herpes Classification
Human herpes virus has the following subfamilies:
1.
Alphaherpesvirinae
simplex virus
HHV-1 = HSV-1: herpes simplex virus-1
HHV-2 = HSV-2: herpes simplex virus-2
varicella virus
HHV-3 = VZV: varicella zoster virus
2.
Betaherpesvirinae
cytomegalovirus
HHV-5 = CMV: cytomegalovirus
roseolovirus
HHV-6
HHV-7 Both cause roseola.
3.
Gammaherpesvirinae
lymphocryptovirus
HHV-4 = EBV: Epstein-Barr virus
rhadinovirus
HHV-8 = KSHV: Kaposi’s sarcoma-associated herpesvirus
6.1.2 HSV and Sudden Deafness
Hondo et al. [1] reported an association between HSV and sudden deafness. They studied the prevalence of HSV-1 in patients with sudden deafness by obtaining serum from patients during the acute phase of hearing loss and two to three times during the recovery phase of the disease. One hundred thirty-eight samples from 49 patients were submitted for neutralization testing (NT) and complement-fixation testing (CFT). Neutralizing antibodies against HSV-1 were present in 38 of 49 patients (78 %), a rate similar to that of the control group (77 %). No significant differences were found among different age groups. The CFT results were positive in 38 of 49 patients (78 %), significantly different from results in the control group, where 54 % of people tested positive. The difference between the NT and CFT results in the control group indicate no reactivation of latent HSV infection after initial infection in some individuals, resulting in the disappearance of CF antibody over time. The high incidence of CF antibodies among the patients with hearing loss suggests that if HSV causes the disease, the mechanism of infection may be related to reactivation of latent infection.
A significantly greater percentage of patients with hearing loss (43 %) had neutralizing antibodies against HSV-2 than the control group (20 %). All patients with antibodies against HSV-2 also had antibodies against HSV-1, suggesting that reinfection with HSV-1 precedes infection with HSV-2.
6.1.3 Morphological Study
6.1.3.1 Experimental HSV Labyrinthitis
In one study, HSV-1 and HSV-2 were isolated from the livers of neonates who died of disseminated HSV infection. The viruses were passaged in human embryonic fibroblasts. Virus titers of the strains were 9.2 × 107 pfu/mL (pfu, plaque-forming units) (type 1) and 3 × 106 pfu/mL (type 2).
Inoculation of the Middle Ear
The middle ear cavities of guinea pigs were inoculated with HSV solution (2 × 103 pfu/2 μL) through a hole in the tympanic bulla or tympanic membrane. Between 2 and 14 days after inoculation, the middle and inner ears were removed, fixed in buffered formalin, and decalcified using 10 % ethylenediamine tetraacetic acid (EDTA) solution. Conventional paraffin wax or celloidin were used for embedding. Viral antigen was detected in the inner ear by indirect immunofluorescence. Anti-HSV antiserum was obtained from rabbits after intracorneal inoculation with the virus and used as the primary antibody. Anti-rabbit IgG conjugated with fluorescein isothiocyanate (FITC) was used as the secondary antibody [2].
Results
HSV types 1 and 2 produced similar pathological changes [3].
A variable number of days after inoculation with the HSV solution, the labyrinths with the bullae were removed and processed. HSV infection of the middle ear caused bleeding, cell infiltration, fibrosis, and new bone formation in the mucosa. Immunofluorescence confirmed the presence of HSV antigen in middle ear infiltrates.
In one animal, the round window was found ruptured on day 11 after inoculation. Massive cell infiltration was observed in the scala tympani of the basal turn after round window rupture. Many multinuclear cells were present in the exudate. Some macrophages were found, each containing 4–5 neutrophils (emperipolesis) (Fig. 6.1) [4]. Inflammation extended to the organ of Corti through the basilar membrane. Exudate was present in the scala tympani of the entire cochlea, with less toward the apex. Mixed infection was considered.
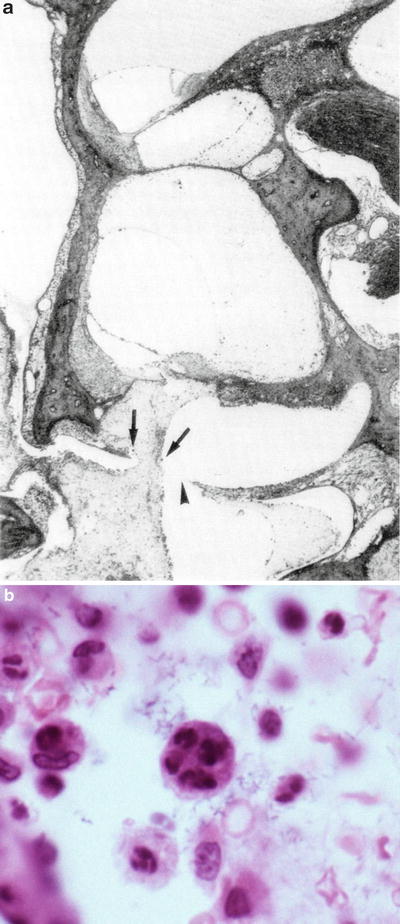
Fig. 6.1
Transtympanic HSV labyrinthitis. (a) Invasion of middle ear infection into the cochlea through the round window membrane. Exudate containing many multinuclear cells (arrows) has reached the scala tympani 11 days after inoculation of HSV-1 into the middle ear. Arrowhead: perforation of the round window membrane (original ×2.5). (b) High power view of the area indicated by the left arrow in (a). A macrophage contains four or five neutrophils (emperipolesis) (original ×40) [4]
Another animal showed marked cell infiltration and hemorrhage from the lower spiral ligament continuously to the scala tympani, and serous inflammation in the scala vestibuli and scala media 10 days after middle ear inoculation. The stria vascularis showed marked edema with disappearance of intermediate cells and was about twice as thick as normal. Edema was also observed in portions of the spiral ligament (Figs. 6.2a, b, 6.3) [4].
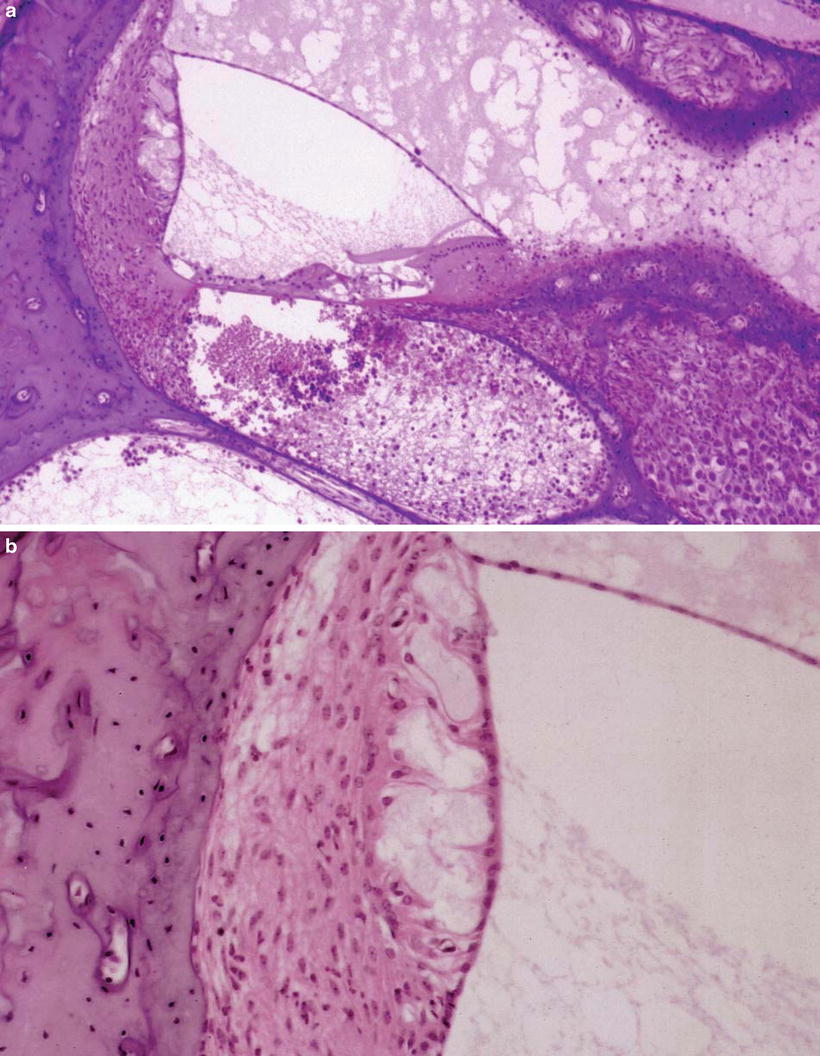
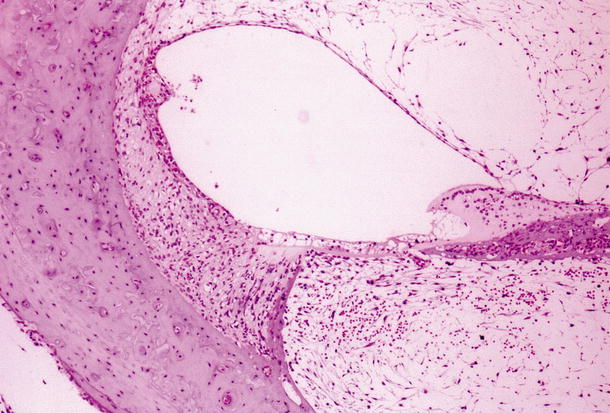
Fig. 6.2
Transtympanic HSV labyrinthitis. (a) Viral labyrinthitis 10 days after HSV-1 inoculation of the middle ear. Inflammation of the cochlea spreads mainly in the scala tympani of the basal turn through the round window. Marked hemorrhage and cell infiltration is observed in the lower spiral ligament and scala tympani. The stria vascularis shows marked edema and loss of the intermediate cells (original ×6.5). (b) Enlarged stria vascularis. Loss of intermediate cells and edema are marked. The spiral ligament is also edematous (original ×16) [4]
Fig. 6.3
Transtympanic HSV labyrinthitis. Seven days post-inoculation of the middle ear. Marked fibrosis in the scala tympani and scala vestibuli of the basal turn. The organ of Corti has degenerated. The stria vascularis is atrophic and has trapped the detached tectorial membrane (original ×6.5)
Invasion of middle ear infection also occurred without rupture of the round window membrane. In such cases, it was hard to distinguish the structure of the round window membrane because of the surrounding marked cell infiltration.
Inoculation of the Scala Tympani
HSV solution (2 × 103 pfu/2 μL) was inoculated directly into the scala tympani through the round window membrane using a fine needle. A small piece of absorbable gelatin was placed over the round window membrane and the skin incision was sutured after inoculation.
The scalae tympani and vestibuli were fibrotic 7 and 9 days after inoculation. This change was marked in the basal turn and less prominent toward the apical direction. Viral antigens were detected in the organ of Corti, Reissner’s membrane, limbus spiralis, stria vascularis, spiral ganglion, and in fibrous tissue in the scalae tympani and vestibuli 6–12 days after virus inoculation into the scala tympani.
The organ of Corti degenerated after HSV inoculation. The nuclei of the hair cells disappeared first, while the cell bodies persisted for a time. Changes in the inner hair cells were less marked than in the outer hair cells. The supporting cells were resistant to infection. There was heavy distribution of antigen in the basal turn where morphological changes were prominent.
Electron microscopy showed HSV nucleocapsids in the nerve fibers and endings of afferent and efferent nerves beneath the outer hair cells (Fig. 6.4) [5]. The nerve fibers in the inner spiral bundle contained HSV nucleocapsids.
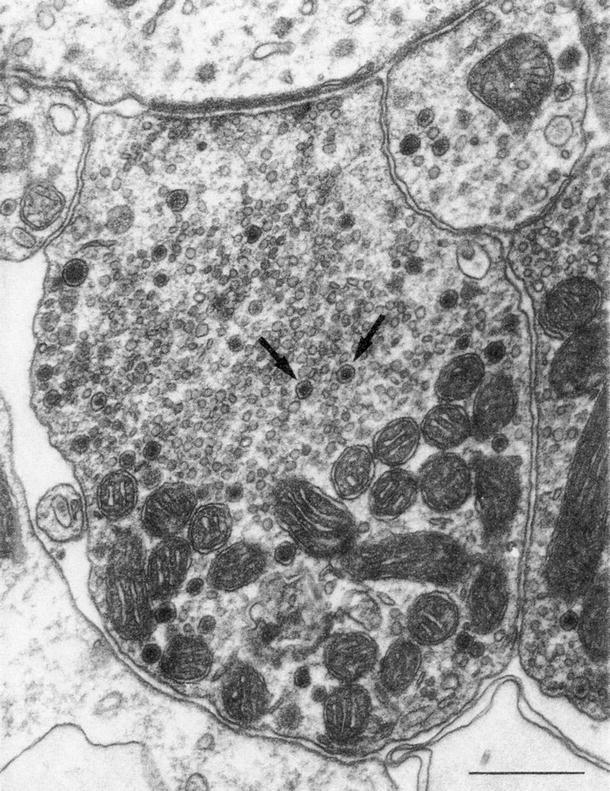
Fig. 6.4
HSV nucleocapsids (arrows) in one of the efferent nerve endings beneath the first row of outer hair cells of the third turn. Scale: 0.5 μm × 31300 [5]
After HSV infection, the tectorial membrane often detached from the limbus spiralis and became rounded or floated within the scala media. Detached tectorial membrane was trapped either by Reissner’s membrane or by the stria vascularis (Figs. 6.4 and 6.5).
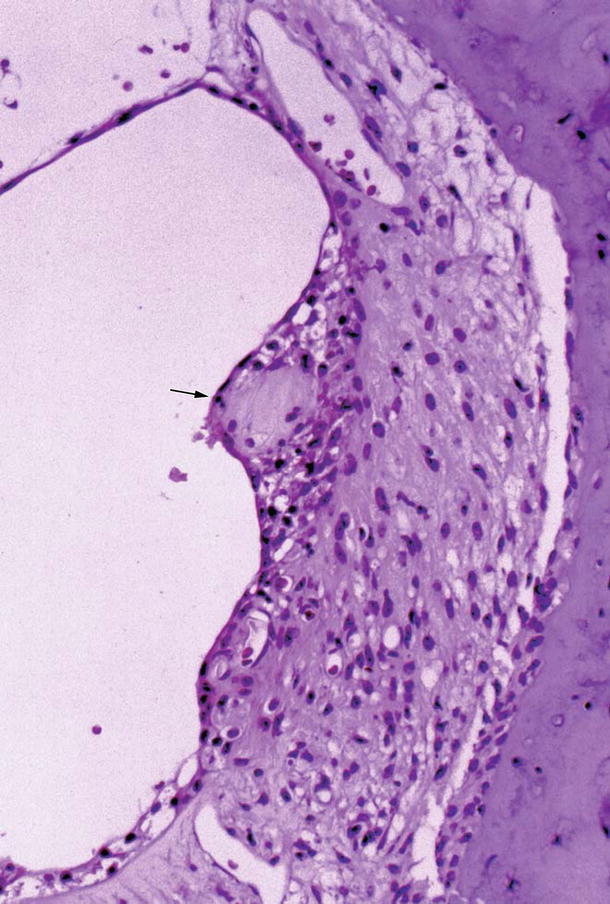
Fig. 6.5
The stria vascularis in viral labyrinthitis 7 days after intracochlear inoculation of HSV-1. The stria vascularis has trapped the detached tectorial membrane, which is covered by atrophic marginal cells. There is cell infiltration around the tectorial membrane. The arrow indicates the tectorial membrane (original ×16)
Sometimes degenerated tectorial membrane became a round mass, attached to Reissner’s membrane, and was encapsulated by the epithelial cell layer of Reissner’s membrane. Similarly, detached tectorial membrane trapped by the stria vascularis was covered by the marginal cells, and disappeared in the stria (Fig. 6.5). Within the stria vascularis, there was cell infiltration around the tectorial membrane. In the stria vascularis the tectorial membrane would be digested and disappear, while tectorial membranes attached to Reissner’s membrane would remain unchanged for a long time.
Immunofluorescence studies revealed the distribution of HSV antigen. Particularly strong fluorescence was found on the surface of rolled-up tectorial membrane. The interdental cells also showed strong fluorescence (Fig. 6.6) [5].
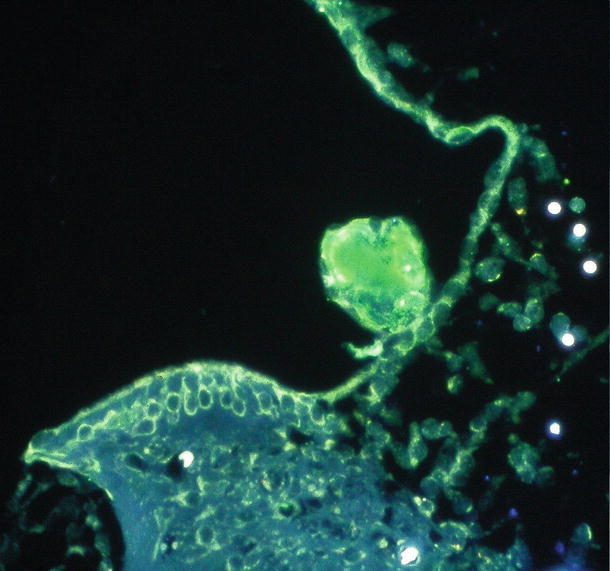
Fig. 6.6
Presence of HSV antigen in the rolled-up tectorial membrane 9 days after intracochlear inoculation. Intense fluorescence reaction in the rolled-up tectorial membrane indicates the presence of HSV antigen. Strong reaction is also found in Reissner’s membrane and in the interdental cells (original ×16) [5]
Another tectorial membrane change was the presence of many small particles on the surface of the marginal and middle zones in the third turn. In HSV labyrinthitis, the matrix of the tectorial membrane showed aggregation of variably-sized masses on the surface, as well as within the tectorial membrane (Figs. 6.7 and 6.8a–c) [3, 6]. Similar findings were observed on the tectorial membrane of a patient who developed acute bilateral profound sensorineural hearing loss over the course of a single day (see text in Chap. 4, Sudden Deafness). Non-enveloped viral particles were found on the surface of the masses or bulges formed on the surface of the tectorial membrane in the experimental animals.
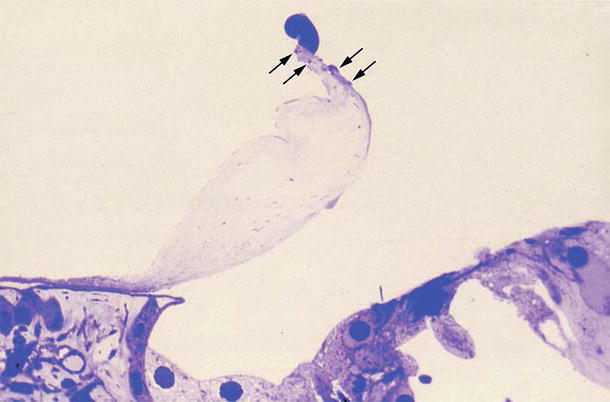
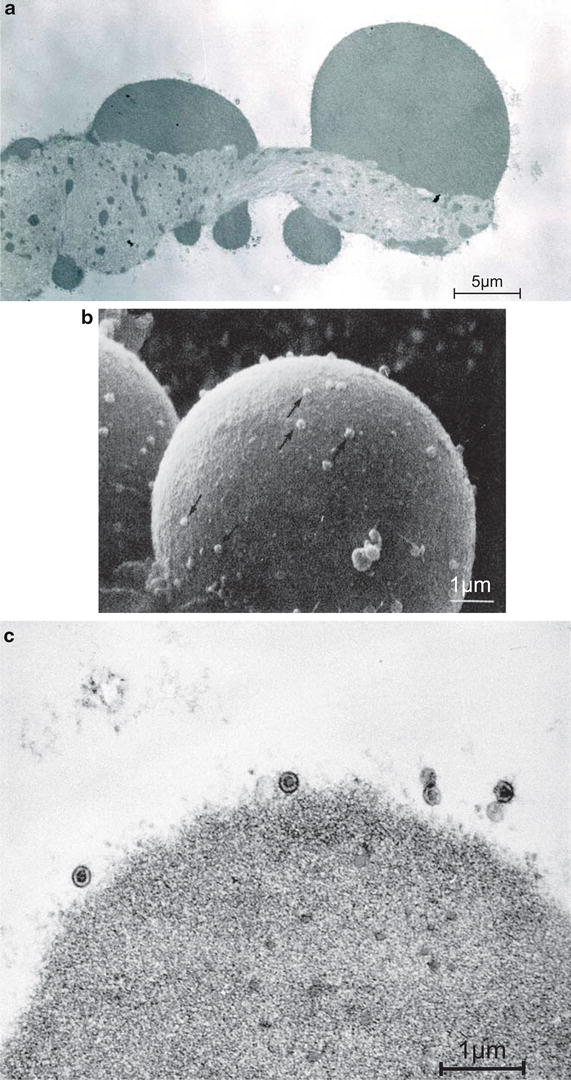
Fig. 6.7
Degeneration of the tectorial membrane. Seven days after intracochlear inoculation with HSV-1. Bulge formation at the tip of the marginal zone. Several small bulges are present nearby (arrows). Loss of interdental cells is observed. Toluidine blue staining (original ×16) [3]
Fig. 6.8
Degenerated tectorial membrane with viral particles. (a) Transmission electron microscopic observation 9 days after intracochlear inoculation with HSV-1. Larger masses are present on the surface of the degenerating tectorial membrane. Many small masses are observed within the membrane (Bar: 5 μm, original ×7,500) [3]. (b) Scanning electron microscopic observation. A spherical mass on the surface of the tectorial membrane consisted of condensation of degenerated filaments. Many viral particles are seen on its surface (arrows) 7 days after intracochlear inoculation (Bar: 1 μm, original ×12,000) [9]. (c) Transmission electron microscopic observation. Viral particles on the surface of a bulge 9 days after intracochlear inoculation (Bar: 1 μm, original ×54,000 [3])
HSV particles consist of three main parts: the core, the characteristic shell (capsid), and the envelope [7]. It is rather unusual for HSV particles in the extracellular space to have no envelope. The unusual finding of non-enveloped virus particles in this case may be due to the presence of endolymph within the extracellular space.
The spiral ganglion cells are the preferred site of HSV infection. In the first days after inoculation, two types of intranuclear inclusion bodies appeared in the spiral ganglion: Cowdry type A and full type. The spiral ganglion cells gradually disappeared [8].
It is noteworthy that viral antigens were detected on the sixth day in the spiral ganglion not only on the inoculated side, but also on the non-inoculated side (Fig. 6.9) [5]. The opposite cochlea showed normal morphology, so the presence of virus there did not result from spread of viral inflammation from the inoculated cochlea. Potential routes of virus migration to the contralateral spiral ganglion include (1) through the CSF, (2) through the perineural space, (3) through the perilymph and CSF, or (4) hematogenous spread. The phenomenon of Schreiner may be involved. In the absence of morphological changes, the presence of HSV in the non-inoculated cochlea would be termed a latent infection. We attempted but failed to reactivate latent infection to induce drastic changes in the cochlea with administration of cyclophosphamide. If viral reactivation becomes possible, this method will be used to produce a model of acute profound sensorineural hearing loss.
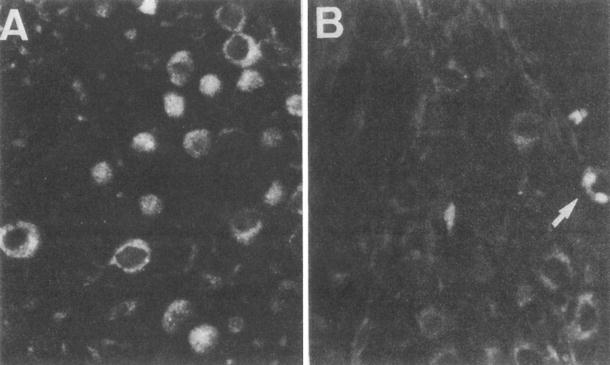
Fig. 6.9
HSV antigen in the spiral ganglion. (a) The inoculated side of the spiral ganglion shows the presence of HSV antigen. (b) The non-inoculated side of the ganglion shows the presence of antigen only sporadically. A single ganglion cell has HSV antigen in this photograph (arrow) (original ×16) [5]
Five to seven days after intralabyrinthine inoculation with HSV-1, the undersurface of the tectorial membrane showed several changes, while the hair cells and supporting cells showed no change.
Imprints exist on the undersurface of the tectorial membrane. Under a scanning electron microscope, partial exposure of the fibrous layer of the tectorial membrane was found after HSV infection, resulting from loss of the membrane matrix. Furthermore, changes were found in the imprint itself. The normal imprint consists of several elliptic dips arranged independently. In the infected tectorial membrane, the dips disintegrate, and may be fused (Fig. 6.10a–c) [9]. The coupling of the stereocilia with the deformed dip will interfere with normal sound transmission.
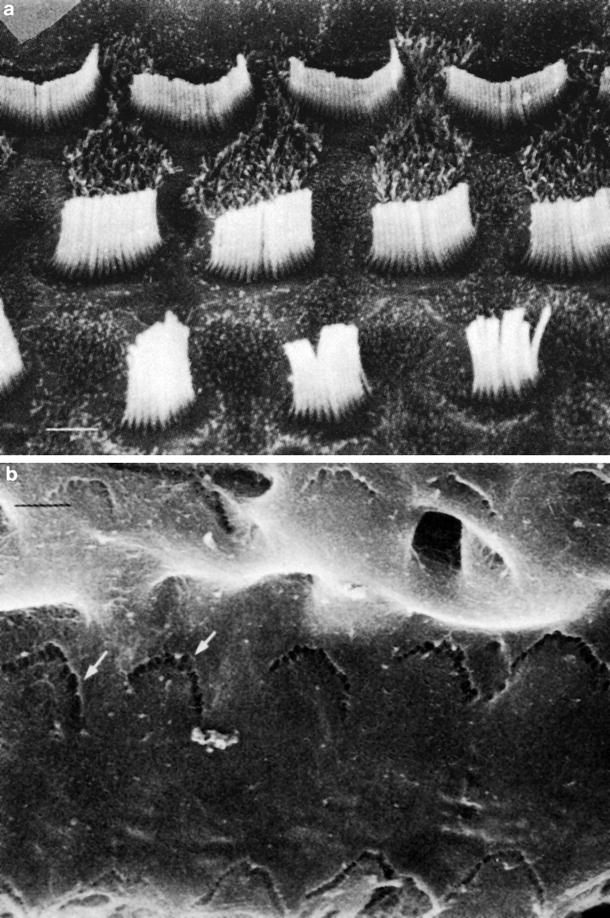
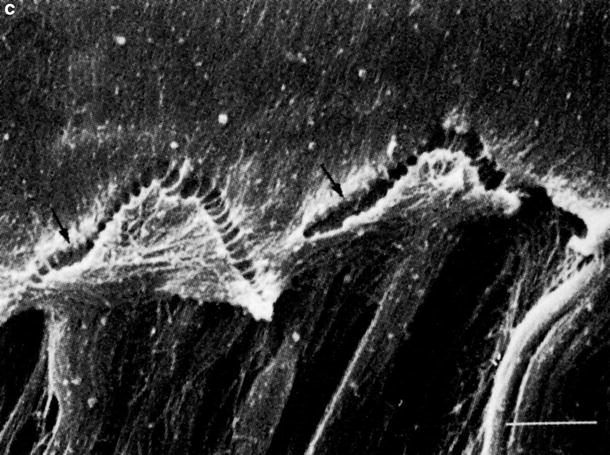
Fig. 6.10
The stereocilia and imprints. Scanning electron microscopic observations 7 days after intracochlear inoculation of HSV-1 (1 × 106 pfu/2μL). (a) Stereocilia of the outer hair cells. There are no apparent changes (Bar: 1 μm). (b) Dips in the imprints are destroyed (arrows) (Bar: 1 μm, original ×20,000). (c) Fusions of the imprint (arrows) and exposure of the fibrous layer (Bar: 1 μm, original ×20,000) [9]
More advanced changes were observed in the cochlea 2 months after intralabyrinthine inoculation. The infected scala tympani revealed fibrosis and ossification. The stria vascularis had disappeared. Marked endolymphatic hydrops had developed. The organ of Corti had a strange appearance, with only the surface remaining as a thin membranous structure. The border cells, pillar cells, membrana reticularis, and Hensen’s cells were probably involved in the formation of the structure. However, no visible cell elements were found inside the organ of Corti. No cellular infiltrates were observed within the cortilymphatic space (Fig. 6.11a, b).
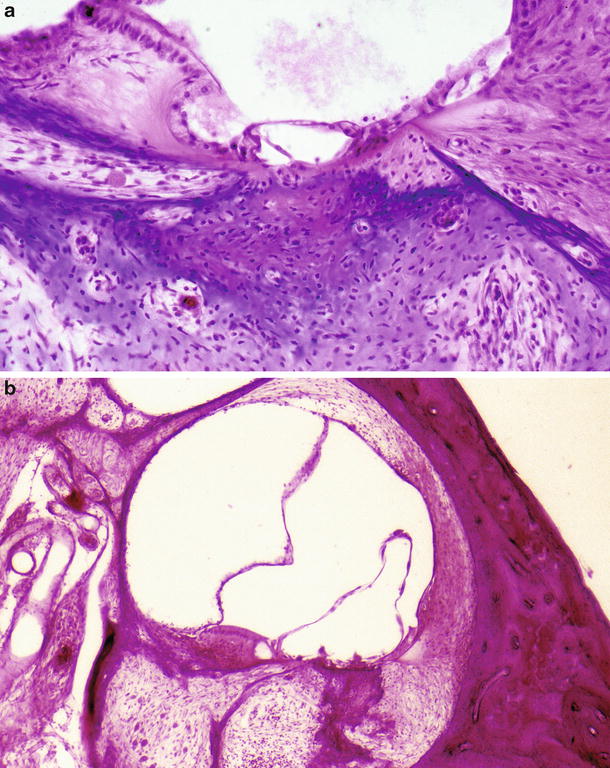
Fig. 6.11
Viral labyrinthitis 2 months after HSV inoculation of the scala tympani of the basal turn. (a) Ossified scala tympani of the basal turn. Surface of the organ of Corti remains. No hair cells or supporting cells remain. There is no cell infiltration within the organ of Corti. Nerve fibers in the osseous spiral lamina are missing (original ×6.5). (b) The organ of Corti has disappeared, leaving its surface as a bulge. The scala media shows endolymphatic hydrops with cortilymphatic hydrops. The scala tympani is filled with fibrous tissue and new bone. The osseous spiral lamina and Rosenthal’s canal are enlarged. Marked loss of the spiral ganglion cells is observed (original ×2.5)
Perhaps apoptosis had taken place in the hair cells and supporting cells. High osmolarity in the cortilymph induces influx of water from the scala tympani, resulting in the formation of cortilymphatic hydrops (see Chap. 2, Inner Ear Fluid).
6.1.3.2 Human Temporal Bone
To my knowledge, HSV labyrinthitis in human temporal bones has not been reported. We know from experiments that HSV infection results in peculiar and characteristic changes in the tectorial membrane. The changes seen with HSV labyrinthitis include rolling up of the tectorial membrane and formation of variably-sized particles resulting from condensation or aggregation of altered tectorial membrane constituents.
The patient with sudden deafness introduced in Chap. 4, in whom viral labyrinthitis was suspected, had tectorial membrane changes quite similar to those observed in experimental HSV labyrinthitis (Fig. 4.4).
Reptiles, a famous Escher lithograph, is suggestive of the viral life cycle. Many sleeping lizards (viruses) are hidden within a white page (a cell) in a drawing book. One of the viruses awakes and escapes from the cell for a walk, becoming gradually more vigorous. Finally, the virus is in high spirits and sits proud of its triumph on a regular dodecahedron (Fig. 6.12).

Fig. 6.12
Reptiles, a lithograph by M.C. Escher printed in 1943 (M.C. Escher’s “Reptiles” © 2013 The M.C. Escher Company – The Netherlands. All rights reserved. www.mcescher.com)
After its excursion, the virus returns to lie dormant in the cell. This illustrates the cycle of reactivation of latent HSV infection, although the nucleocapsid of HSV is a regular icosahedron.
6.2 Cytomegalovirus
6.2.1 Introduction
Cytomegalovirus (CMV) belongs to the Betaherpesvirinae subfamily of Herpesviridae. It is also a member of the TORCH complex, i.e., Toxoplasmosis, Others, Rubella, CMV, and Herpes simplex virus. When these microorganisms infect pregnant women who lack immunity, severe complications may occur in the children before or after birth, without apparent symptoms in their mothers.
Cytomegalic inclusion diseases have been considered a leading cause of human congenital viral infection and hearing loss, including a progressive type of hearing loss in children [10]. According to Barbi et al. [11], more than 40 % of cases of deafness of unknown cause that need rehabilitation are attributable to CMV, which is most commonly the result of intrauterine infection. Infants can also be exposed to CMV through breast milk. Delayed onset of hearing loss after CMV infection is common. Infants with CMV and normal hearing at birth should be monitored for 6 years for hearing loss. Newborn infants with CMV can be treated with ganciclovir.
6.2.2 Incidence of Congenital CMV Infection Among Newborns in Japan
Because congenital CMV infection causes significant clinical consequences not only at birth but also with delayed otological and neurological sequelae, it is critical to establish a strategy for screening congenitally infected newborns. Koyano et al. [12] have recently developed a quantitative PCR assay using urine filter discs placed in diapers. The discs function directly as a PCR template without additional steps. Using these discs, the researchers established a multicentric congenital CMV screening program.
More than 21,000 newborns were enrolled at 25 sites in geographically separate areas of Japan. Congenital CMV infection was identified in 0.31 % (95 % CI, 0.24 % to 0.39 %) of newborns. Thirty percent of these cases (20/66) had typical clinical manifestations and/or had pathological brain imaging results at birth. Audiological testing was conducted using auditory brainstem responses and auditory steady-state responses, and brain imaging was performed using CT and/or MRI. Almost two-thirds of the cases had siblings, a significantly higher rate than for uninfected newborns. When tested, most of these (21/25) were found to have CMV strains identical to those of their siblings. Siblings are a major risk factor for congenital CMV infection.
Ogawa et al. [13] tested 67 children with severe sensorineural hearing loss for CMV and GJB2 mutations. DNA specimens were prepared from dried umbilical cords. Congenital CMV infection was identified in 15 % of the patients, and GJB2 mutations were identified in 24 %. All CMV-positive children had developed sensorineural hearing loss before the age of 2 years. Most had no obvious clinical abnormality at birth.
6.2.3 Histopathology of the Temporal Bone in a Case of CMV
Myers and Stool [14] reported the case of a 3-day-old girl who died of cytomegalic inclusion disease that included exfoliative dermatitis, hepatosplenomegaly secondary to extramedullary hematopoiesis, biliary cirrhosis, and severe bronchopneumonia. Autopsy revealed cytomegalic inclusions in the convoluted tubules of the kidneys and many other organs. Inclusion-bearing cells were also seen in the temporal bones.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


