Fig. 31.1
(a) Example rate vs ILD function. Distributions of half-max ILD (b), ILD dynamic range (c), and rate-ILD slope (d) for low (<4 kHz)- and high (>4 kHz)-frequency ICC neurons
2.2 Acoustical Measurements of ILD as a Function of Source Distance
ILDs were calculated from acoustical measurements made at different sound source-to-observer distances (10, 20, and 100 cm). Recordings took place in a double-walled, acoustic chamber (see Koka et al. 2011 for details). A cone-shaped speaker (MF1-M, TDT, Alachua, FL), creating an acoustic point source, was attached to an industrial sliding camera tripod so that the source could be placed precisely at the required distances from the animal. The animal was placed on a platform such that it could be rotated around its interaural axis through ±90° in 10° steps. The free-field-to-eardrum acoustic impulse response for each ear and each location was measured from which head-related transfer functions (HRTFs) were computed. The acoustic ILDs at each position were calculated by subtracting the left ear HRTFs from right ear.
3 Results
3.1 Low-Frequency ICC Neurons Code for ILDs Outside the Acoustic Physiological Range
Physiological data were based on 103 ILD-sensitive ICC neurons. Figure 31.2 plots the neuronal half-max ILD (symbol) plus the ILD dynamic range (error bars) for each neuron as a function of CF. The dynamic range indicates the ILDs over which the neural response was modulated, while the half-max ILD gives the midpoint ILD about which the response is modulated maximally (Fig. 31.1a). The black line in Fig. 31.2 indicates the maximum acoustical ILD measured at a 1 m (100 cm) distance as a function of frequency computed across all locations in the frontal hemisphere. Consistent with the Duplex theory, ILDs are small (<∼1–3 dB) until the frequency exceeds 4 kHz.
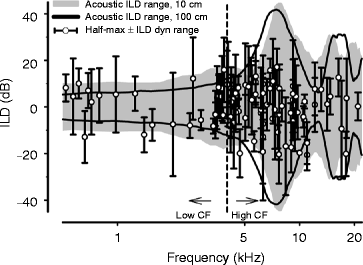
Fig. 31.2
Maximum acoustic ILDs (100 cm, black line) and ICC neuron coding as a function of frequency. Half-max ILD (○) and dynamic range (error bars) are shown for each ICC neuron. Shaded area indicates max acoustic ILDs for source distances 10 cm from the head
Consistent with the acoustics, Heffner et al. (1994) demonstrated that chinchillas use ILDs to localize free-field sounds above 4 kHz and likely use ITDs below 4 kHz. Based on these empirical behavioral and acoustics data, our neurons were separated into low (<4 kHz) and high (>4 kHz) CF groups (vertical dashed line, Fig. 31.2). Two points are observed from Fig. 31.2. First, the distributions of the ILD sensitivity metrics (e.g., Fig. 31.1a) for neurons with high CFs fall within (half-max ILD) and generally span (ILD dynamic range) the range of the empirically measured ILDs available for far-field sources (100 cm; solid black lines, Fig. 31.2). The majority of the half-max ILD values (95 %) for high-CF neurons (69/73) fell within the range of maximum ILDs.
On the other hand, the half-max ILDs of the majority (∼60 %, 18/30) of low CF neurons (<4 kHz) fell at the edge (within 1 dB) or outside of the acoustic range of ILDs. That is, the neural sensitivities of low-frequency ICC neurons were not constrained to the physiological range of ILDs. To test the hypothesis that low-frequency neurons might have different ILD coding capabilities than high-frequency neurons, the distributions of half-max ILD, ILD dynamic range, and the rate-ILD slope (Figs. 31.1b–d) were examined. No significant frequency-dependent differences between the means were observed for any of the ILD coding parameters . These data suggest that the coding of ILDs by ICC neurons is frequency independent and that the low-frequency neurons share essentially identical sensitivities to the same overall ranges of ILDs as high-frequency neurons.
Given a strict interpretation of the Duplex theory, the discrepancy between the frequency independence of neural ILD coding (Fig. 31.1b–d) and frequency dependence of the acoustic ILDs is unexpected as it suggests that there are neurons that can encode sound features that may not be experienced in the natural environment. Alternatively, we suggest that this discrepancy arises from an inadequate characterization of the acoustical environment. Based on prior studies (Duda and Martens 1998; Brungart and Rabinowitz 1999; Kim et al. 2010), we hypothesize that low-frequency ILD-sensitive neurons are necessary because acoustically low-frequency ILDs become large and potentially useful when sound sources are located near the head.
3.2 The Effects of Sound Source Distance on the ILD Cues to Location
Acoustic ILDs were measured as a function of azimuth and three source distances (10, 20, and 100 cm) in three chinchillas. These findings are summarized in Fig. 31.3 where the ILDs are plotted at one azimuth, 75°, as a function of frequency for the three different source distances in each animal – the trends at this source location are representative of other locations. ILD magnitudes generally increased with (1) increasing frequency, (2) with more lateral azimuths away from midline, and (3) with decreasing source distance. At 100 cm (far field), maximum ILDs were very small for low frequencies but become systematically larger for frequencies above ∼4 kHz. From ∼6 to 18 kHz, the spectral notches created by the contralateral pinna (farthest from the source) produce large ILDs. For sources very close (10 cm) to the head, there was a substantial increase (10 dB or more) in ILD magnitude particularly for lower frequencies (<4 kHz). These data demonstrate that ILDs are not only a joint function of azimuth and frequency but also sound source distance.
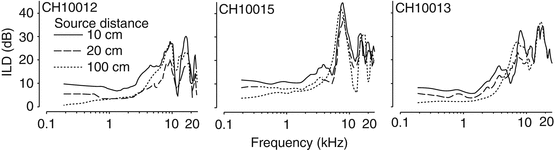
Fig. 31.3
Acoustic ILD measurements at 75° azimuth for three source distances in three animals
3.3 ILD Sensitivity of ICC Neurons Spans the Range of Physically Available Acoustic ILDs when Source Distance Is Considered
The neuronal half-max ILD (symbol) plus the associated ILD dynamic range (error bars) for each neuron were plotted again as a function of the neuron’s CF and then compared to the maximum ILDs that were recorded at distances of 10 cm (gray-shaded area, Fig. 31.2). After taking into consideration the ecologically relevant effect of source distance on ILDs, the majority (93 %) of low-frequency ILD-sensitive neurons (28/30, see Fig. 2 below the 4 kHz dashed line) now have half-max ILD values that fall within the range of available ILDs. As seen in Fig. 31.2, ILDs systematically increase with decreases in the distance between a sound source and observer, particularly for frequencies <∼4–5 kHz. There were still some low-frequency half-max values (2/30) that fell just outside the acoustic ILDs; however, it is expected that for sound source distances <10 cm (the closest distance used here), the maximum ILDs available would be considerably larger, up to 20 dB (Duda and Martens 1998). Thus, the half-max ILDs of these outliers would then fall into the acoustical range of ILDs experienced for sources occupying space close to the head.
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


